 |
 |
|
| Recentemente,
um estudo de DNA publicado por oito autores entre os quais eu (van den Berg
et al., 2000) confirmou algo que há muito se dizia na literatura,
de que as espécies de Laelia ocorrentes no Brasil na realidade
não pertencem ao mesmo grupo das espécies do México.
Devido ao fato de que a primeira espécie a ser descrita no gênero (a espécie-tipo Laelia speciosa) era do grupo das mexicanas, a única alternativa nomenclatural envolvia re-acomodar as espécies brasileiras em outros grupos. |
 |
|
Em um artigo publicado no mesmo número do periódico em que
publicamos o estudo de DNA, publicamos a nossa alternativa de transferir
as espécies brasileiras de Laelia para Sophronitis
e todas as combinações necessárias (van den Berg e
Chase, 2000). Recentement, Chiron e Castro Neto (2002) publicaram uma outra alternativa taxonômica, que envolveu distribuir estas espécies por diversos gêneros, três dos quais novos: Dungsia, Hadrolaelia, Hoffmannseggella, Microlaelia, além de Sophronitis. Nas considerações a seguir, pretendo discutir as razões que nos levaram a agrupar todas as espécies em Sophronitis e fazer comentários sobre as diferentes questões científicas envolvidas neste assunto. Vale frisar que não existe em momento nenhum qualquer questão pessoal entre mim e Chiron e Castro Neto, com quem mantenho relações cordiais. Quando tratamos de questões de classificações, embora o relacionamento filogenético entre um grupo de espécies (as árvores mostrando os relacionamentos de parentesco) possa ser estabelecido de forma objetiva (embora não obrigatoriamente correta), as classificações resultantes (os nomes que serão aplicados) tem um caráter subjetivo, já que temos que escolher a que grupos serão atribuídos níveis hierárquicos (seções, subgêneros, gêneros, subtribos, tribos, famílias e assim por diante). Os dois extremos óbvios desta escolha seriam de juntar ao máximo um grupo (por exemplo, juntar todas as Cattleyas, Sophronitis, Brassavola em um só gênero = Brassavola), ou dividir de tal maneira que cada espécie destes gêneros seria colocada em um gênero com uma única espécie e teríamos centenas de gêneros. |
| Níveis
intermediários entre estes extremos visam maximizar a informação
filogenética, isto é, ao agruparmos um certo número
de espécies dentro de um nível, por exemplo, gênero,
estamos retendo a informação de que aquele grupo de espécies
é mais relacionado entre si do que com outros grupos. Com o avanço dos estudos de sistemática molecular em plantas, e uma conseqüente reorganização na maioria dos grupos (não é só nas Sophronitis que isto está ocorrendo), uma série grande de rearranjos começaram a ser feitos e começou-se a tentar estabelecer alguns critérios para a redelimitação de grupos. Muitos desses foram resumidos por Backlund e Bremer (1998) e também utilizados na nova classificação de todas as plantas com flores (Angiosperm Phylogeny Group, 1998): 1) Evitar grupos monotípicos (os que contém apenas uma espécie). Grupos com uma única espécie representam a perda total de informação, já pelo nome não é possível associar aquela espécie com nenhuma outra. 2) Procurar alterar na medida mínima do possível a nomenclatura existente. Neste sentido, as mudanças nomenclaturais devem ser feitas apenas quando necessárias (ex. porque um determinado grupo não pode ser mantido). De forma similar, os grupos propostos devem procurar ser estáveis (com menor chance de mudança) e, na medida do possível, conservarmos o tamanho dos grupos. |
Quando
escolhemos transferir as espécies brasileiras para Sophronitis,
vários fatores foram levados em consideração: |
 |
| No sistema de Chiron e Castro Neto (2002) espécies bem constrastantes
como Sophronitis purpurata e Sophronitis brevipedunculata continuam sendo parte do mesmo gênero. |
| |
 |
| A
segunda argumentação é de que o nosso sistema ignora
o de nossos predecessores. Novamente uma afirmação infundada,
já que toda a taxonomia infra-específica que existia em Laelia
poderia ser passada para Sophronitis, embora preferimos deixar este
passo para fazer de forma mais criteriosa com mais dados de DNA. Por outro
lado, ao criar gêneros para estas categorias inferiores, Chiron e
Castro (2002) deram um passo arriscado em relação aos sistemas
inferiores, mudando-as de nível hierárquico. Do ponto de vista
morfológico não parece haver também suporte na literatura
para separação em gêneros menores. O Dr. Soto Arenas,
que fez a única outra análise que existe sobre filogenia de
Laelia, esta com base em dados morfológicos (Halbinger e Soto
Arenas, 1997), foi um dos co-autores em nosso trabalho de DNA e seguidamente
discutimos a colocação em Sophronitis antes de tomarmos
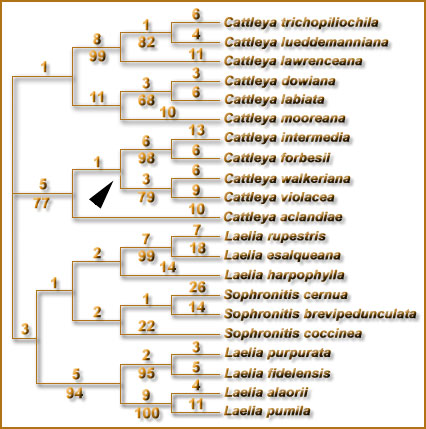
esta decisão. No trabalho de Chiron e Castro (2002) embora seja dada uma justificativa morfológica, uma leitura atenciosa do trabalho deixa claro que todos os grupos foram delimitados exclusivamente com base nas árvores de van den Berg et al. (2000), da mesma forma que a lista de espécies não difere da lista publicada em Sophronitis no artigo adjacente. Nenhum dado adicional foi apresentado para justificar as decisões. Isto nos preocupou sobremaneira pois conhecemos a fragilidade dos dados de ITS para estabelecer grupos dentro de Sophronitis (que foi expressa seguidamente no trabalho original). Isto é ainda mais surpreendente pois no início do trabalho Chiron e Castro (2002) citaram cinco considerações preliminares sobre utilização de árvores filogenéticas para classificação (ítens A-E), para, logo em seguida, desrespeitar estes critérios quase imediatamente (especialmente o ítem E sobre a robustez dos ramos). Embora ainda não estejam concluídos novos estudos que estamos fazendo com várias outras regiões de DNA para verificar as relações dentro de Sophronitis, dados preliminares com duas regiões dos cloroplastos (Fig. 1) já demonstram que para utilizar o sistema de Chiron e Castro (2002), que acaba de ser proposto, modificações substanciais seriam necessárias. |
 Dados de sequência de plastídeo recentes mostram que S. harpophylla é estreitamente relacionada ao grupo das rupícolas |
Com
base em dados das regiões trnL-F e matK as espécies de Sophronitis
no sentido antigo(antes de receber as espécies de Laelia)
constituem um bom grupo (diferente da separação de S. cernua
que havia na árvore de ITS) e, portanto, a colocação
do grupo de S. coccinea em Hadrolaelia é inadequado.
Da mesma forma, S. harpophylla vai como irmã das rupícolas e novamente surge um impasse. Ao mesmo tempo que as espécies de Dungsia têm algumas diferenças em relação às rupícolas,é inegável que elas compartilham várias outras |
|
. características morfológicas com este grupo e assim
os gêneros Dungsia e Hoffmanseggella teriam que ser
fundidos. Com essas evidências, a única justificativa para termos os dois gêneros separados seria a vontade dos autores. Por todo o exposto acima, voltamos a frisar que a utilização desses pequenos grupos como gêneros é precipitada e muito pouco estável. Ainda, por alguma razão misteriosa, em nenhum lugar do trabalho de Chiron e Castro estes autores mencionaram que havia uma classificação alternativa destas espécies em Sophronitis. Embora isto possa ter sido uma mera distração, o resultado foi pouco ético no sentido de não dar ao leitor a opção de buscar a literatura e conhecer todas as propostas alternativas, conduzindo o leitor a uma alternativa única e como se fosse absoluta novidade. Em entrevista recente à revista “Brasil Orquídeas” o Sr. Chiron disse que não incluiu como sinônimos as Sophronitis para não “chamar a atenção” para os nomes em Sophronitis. Isto novamente soa como se, por alguma razão misteriosa, os autores estivessem omitindo qualquer alternativa à classificação apresentada. Dentro da proposta deste trabalho, queremos ressaltar que a discussão aberta das diferentes idéias é saudável no meio científico e as justificativas científicas para tomarmos uma ou outra decisão taxonômica devem sempre ser feitas de forma explícita. Só assim podemos fundamentar nossas opiniões, e mais importante ainda, dar ao leitor a chance de buscar na literatura e escolher a alternativa que mais lhe convenha. |
 |
|
Fig.
1 - Uma das árvores produzidas em análise de parcimônia
de dados de sequência das regiões trnL-F e matK dos cloroplastos,
para Sophronitis, Cattleya e gêneros próximos. |
Literatura
Citada: |
| Cássio
van den Berg Depto. de Ciências Biológicas Universidade Estadual de Feira de Santana BR 116 Km 3 Feira de Santana, BA 44031-460 Brasil |
Dr.
Cássio van den Berg é agrônomo pela Universidade de
São Paulo, Mestre em Ecologia pela UNICAMP e doutor em Botânica
pelo Royal Botanic Gardens, Kew e The University of Reading (Reino Unido). O seu trabalho com as Laelias brasileiras foi parte das exigências para obtenção do título de Doutor. Todos os trabalhos do autor incluindo o sobre as Laelias brasileiras, podem ser obtidos em sua página web: http://www.cassiovandenberg.hpg.com.br |
| Fotos: Cássio van den Berg ©Cássio van den Berg 2003 |
| Expressamente
proibido qualquer tipo de uso, de qualquer
material deste site (texto, fotos, imagens, lay-out e outros), sem a expressa
autorização de seus autores. Qualquer solicitação
ou informação pelo e-mail bo@sergioaraujo.com |
|
|
voltar |