 |
 |
|
| Recently,
a DNA study published by eight authors including myself (van den Berg et
al., 2000) confirmed a pattern previously mentioned in the literature, that
the species of Laelia occurring in Brazil in reality do not belong
to the same group of the Mexican species. Due to the fact that the first species to be described in the genus (Laelia speciosa) was from the Mexican groups, the only nomenclatural alternative involved to re-accommodate the Brazilian species in some other group or groups. |
 |
|
In another article published in the same journal, we published our choice,
which was to transfer these Brazilian species of Laelia to Sophronitis,
and all necessary combinations were proposed therein (van den Berg and Chase,
2000). Recently, Chiron and Castro Neto (2002) published a different taxonomic choice, which consisted of distributing these species in several genera: Dungsia (new), Hadrolaelia (new), Hoffmansegella, Microlaelia (new), in addition to Sophronitis. In the following lines, I intend to discuss the reasons that have lead us to group all species in Sophronitis and make comments about the different scientific aspects involved in this matter. It is noteworthy to say that there is no personal question between myself and Chiron or Castro, with whom I keep respectful social relations. When we enter into classification matters, although the phylogenetic relationships among a group of species (the trees showing the ancestry relationships) can be established in a more or less objective manner (although not necessarily correct), the resulting classifications (the names which will be used) have a subjective character. This is due to the fact that we still have to choose which groups will be used to establish each hierarchical level (sections, subgenera, genera, subtribes, tribes, families, and so on). Two obvious extremes in such classifications would be to lump large numbers of species in a group (e.g., to lump all Cattleyas, Sophronitis and Brassavola in a single genus = Brassavola), or to split into small groups in such a manner that every genus would include a single species, and we would have hundreds of genera. |
| Intermediate
levels between these two extremes aim to maximize phylogenetic information,
that is, when we group a number of species within a group, we keep the information
that such a group of species is more among themselves than with other groups. With the advance of molecular systematic in plants, and consequently a large amount of reorganization in many groups (it's not only in Sophronitis this thing is happening!), a large number of changes started to be made, and researchers had to propose some criteria for re-delimiting groups. Many of these were summarized by Backlund and Bremer (1998), and others used in the new classification of all flowering plant families in orders (Angiosperm Phylogeny Group-APG, 1998): 1) To avoid monotypic groups (those which contain a single species): groups with a single species represent a total lack of information, since by its name it is not possible to associate that species with any other, 2) To try, inasmuch as possible, not to change the current nomenclature. In this sense, nomenclatural changes should be made only when necessary (e.g. because a group cannot be kept as it is like Laelia). In a similar manner, the groups proposed should try to be stable (with a smaller chance of being changed later), and, if possible, having a similar size to the one before. |
|
When
we chose to transfer the Brazilian species to Sophronitis several
of these factors were taken into consideration: |
 |
| In
Chiron and Castro Neto's system (2002) species really contrasting such as
Sophronitis purpurata and Sophronitis brevipedunculata remain as part of the same genus. |
| |
 |
| The
second statement is that our system ignores that of our predecessors. Again,
this has no basis, since all the infraspecific taxonomy of Laelia
can be transferred to Sophronitis, although we prefer not to do so
without assessing it first with DNA data. On the other hand, creating genera
for all these lower categories, Chiron and Castro (2002) make a risky move
in relation to previous systems, by changing the hierarchical levels. On
a morphological viewpoint also there seems to be no support in the literature
for the separation into smaller genera. Dr. Soto Arenas, which made the
only other analysis that exist on the phylogeny of Laelia, with base on
morphological data (Halbinger and Soto Arenas, 1997), was one of the co-authors
in our DNA study, and we discussed for a long time the placement in Sophronitis
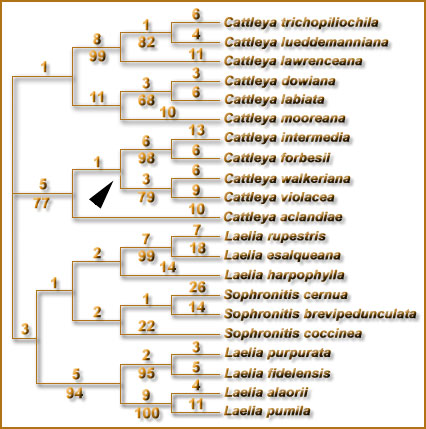
before taking this decision. In the work of Chiron and Castro (2002), although a morphological justification is given, a careful reading makes it clear that all groups were delimited exclusively based on the trees of van den Berg et al., (2000), which were reproduced therein, and also the list of species does not differ from the list published in Sophronitis in our adjacent paper. No additional data was presented to justify the decisions. This somehow made us to worry, because we knew the fragility of ITS data to establish groups within Sophronitis (this fragility was stressed in the original study of DNA). This is even more surprising because in the beginning of their article, Chiron and Castro (2002) cited several preliminary cautions about the use of phylogenetic trees for establishing classifications (items A-E), and immediately ignored these criteria (especially item E about robustness of branches) in their proposal. Although studies with several other DNA regions for assessing relationships within Sophronitis are not finished, preliminary data with two plastid regions (Fig. 1) already show that in order to use the system of Chiron and Castro (2002), which was just proposed, several substantial modifications would be necessary. Based on data from the regions trnL-F and matK, the species of Sophronitis |
 Recent plastid DNA data show that S. harpophylla is closely related to the rupicolous species. |
in the old sense (before receiving the species of Laelia), constitute
a good group (differently from the separation of S. cernua which
appeared in the ITS trees), and therefore the placement of the group of
S. coccinea in Hadrolaelia is inadequate. Similarly, S. harpophylla goes as sister to the rupicolous species, what makes a dubious situation. At the same time that these species placed in Dungsia have some differences in relation to the rupicolous species (especially size), it is obvious that they share many othermorphological characters with |
|
this group, and in this sense it appears better than Dungsia
would be lumped to Hoffmannseggella in their system. In this case the only real justification that could be made to keep Dungsia separated from Hoffmannseggella would be the will of the authors. Based on all facts exposed above, we stress that the use of these small groups of species as genera is hasty and little stable. Still, for some strange reason, nowhere in the work of Chiron and Castro these authors mentioned that an alternative classification of these species in Sophronitis existed. Although this could have been a distraction, the result was a little unethical for not giving the reader the option to look in the literature and knowing alternative proposals, instead misleading the reader to think this was the only solution, and absolute novelty. In a recent interview to "Brazil Orquideas", Mr. Chiron stated that he did not include Sophronitis combinations as synonyms to avoid calling attention to the names in Sophronitis. This again sounds as if the authors were avoiding people to know of any alternative to the classification they presented. In the proposal of this article, we want to reinforce that a discussion of different ideas is a good thing in the scientific world, and that the scientific justifications for taking one or other taxonomic decision should be always made in an explicit way. This is the only way of strengthening our opinions, and even more important, to give the reader the chance to choose in the literature the alternative which is more convenient. |
 |
|
Fig.
1: One of the trees found in a parsimony analysis based on sequence
data from the plastid regions trnL-F and matK, for Sophronitis and related
genera. |
|
Literature
Cited: |
| Cassio
van den Berg Depto. de Ciências Biológicas Universidade Estadual de Feira de Santana BR 116 Km 3 Feira de Santana, BA 44031-460 Brasil |
Dr.
Cassio van den Berg is agronomist, graduated by the Universidade
de São Paulo, Master in Ecology by UNICAMP and Ph.D. in Botany by
Royal Botanic Gardens, Kew and The University of Reading (United Kingdom). His work with Brazilian Laelia has been part of the exigences for obtaining his Phd title. All author's works including the work about Brazilian Laelia can be accessed in his web page: http://www.cassiovandenberg.hpg.com.br |
| All photos by Cassio van den Berg ©Cassio van den Berg 2003 |
|
Any
kind of reproduction (print, digital or anyone other) of any type of
material of this site - texts, layout, photos, images and others - is
strictly forbidden without the previous written permission of the authors. Any solicitation or information should be done by the e-mail bo@sergioaraujo.com |
|
|
|
back |