
|
por
João
A. N. Batista (1), Luciano de Bem Bianchetti(1) & Eduardo G. Gonçalves(2) |
| (1) Embrapa Recursos Genéticos
e Biotecnologia, Parque Estação Biológica, Final Av. W5 Norte, CP 02372, Brasília-DF, 70770-901, Brasil janb@cenargen.embrapa.br bianchet@cenargen.embrapa.br (2) Universidade de São Paulo, Dept. de Botânica, CP 11461, São Paulo-SP, 05422-970, Brasil edggon@hotmail.com |
| Trabalho publicado originalmente na Novon, revista de nomenclatura botânica do Missouri Botanical Garden, volume 13, número 4, páginas 397 a 402, em 2003. |
ABSTRACT.
Habenaria pabstii, a new species (Orchideae, Orchidaceae) from the core region of the cerrado vegetation of central Brazil, is described and illustrated. The species has been collected since 1961, but was repeatedly misidentified as other taxa. The species is similar to those in section Macroceratitae, but distinct by the shorter, forward arched, free spur and the shorter, non-involute stigmas. |
RESUMO.
Habenaria pabstii, uma nova espécie (Orchideae, Orchidaceae) da região nuclear do cerrado do Brasil central é descrita e ilustrada. A espécie tem sido coletada desde 1961, mas foi repetidamente identificada erradamente. A espécie é similar àquelas da seção Macroceratitae, mas distinta pelo calcar mais curto, livre, projetado para frente e pelos estigmas mais curtos e não involutos. |
| Palavras chave: Habenaria, nova espécie, cerrado, Brasil. |
Habenaria
é o segundo maior gênero de orquídeas do Brasil,
compreendendo cerca de 165 espécies de acordo com o ultimo grande
trabalho das orquídeas brasileiras (Pabst & Dungs, 1975).
O gênero é formado exclusivamente por espécies terrestres,
sendo típico de ambientes campestres. A maior parte das espécies
tem flores pequenas e inconspícuas, com poucos atrativos para
os colecionadores sendo, por isto, raramente vistas em cultivo. Como
conseqüência o trabalho com o gênero tem se baseado
quase exclusivamente em coletas botânicas e no exame de material
herborizado. |
 |
Até o momento, os três principais tratamentos das espécies
brasileiras são os de Cogniaux (1893) na Flora Brasiliensis,
Hoehne (1940) na Flora Brasilica e Pabst & Dungs (1975, 1977)
no Orchidaceae Brasilienses. O grande número, a má
caracterização ou pouca informação e a existência
de vários complexos de espécies, resultou numa situação
onde a identificação no gênero é geralmente
complicada, com diferentes nomes sendo aplicados a uma mesma espécie
e um mesmo nome sendo usado para várias espécies diferentes. No Brasil, o principal centro de diversidade de Habenaria é o cerrado, localizado na região central do país. O cerrado é uma savana com alta diversidade de espécies que cobre dois milhões de km² do Brasil Central (Ratter et al. 1997). Uma das áreas, no Brasil Central, mais ricas em espécies é a região nuclear do bioma, incluindo o Distrito Federal, com cerca de 77 táxons (Batista & Bianchetti, 2003) e áreas vizinhas no estado de Goiás. No curso de um inventário florístico das orquídeas desta região foi coletada uma espécie distinta de todas as outras conhecidas do continente Americano. Essa espécie, com flores grandes e ornamentais, é descrita aqui como nova. |
Habenaria
pabstii J.A.N.Bat. & Bianchetti, sp. nov. TYPUS: Brasil. Distrito
Federal: Brasília, Fazenda Água Limpa (UnB), 15o57´S-47o55´W,
1070 m, Córrego da Onça, 2 Fev. 1994, L.B. Bianchetti,
J.A.N Batista & B.M.T. Walter 1489 (holótipo, CEN; isótipos,
HB, K, MO, SP). |
Habenaria
pabstii florum generali aspectu speciebus sect. Macroceratitae Kraenzlin
similis sed spora breviori, libera, frontaliter arcuata et stigmatibus
brevioribus, non involutis differt. Etiam H. urbaniana Cogniaux
similis sed floribus frequenter longioribus et structura columnae prominenti
lobo mediano rostelli praedito appendicem similitudine cavernae formans,
haec ultra antheras producta differt. |
Erva
terrestre. Túbera basal, oblonga a subesférica, 1.2-2.8
x 0.8-1.7 cm; raízes ca. 1 mm largura, esparsamente pilosas,
esbranquiçadas; caule ereto, folioso, 20-60 x 0.3-0.4
cm incluindo a inflorescência. Folhas 7-11, as 3-4 inferiores
em forma de bainhas, as maiores no centro do caule, (3)4-5(6), patentes,
linear-lanceoladas, 11-21 x 1-1.4 cm, agudas, as 1(2) folhas apicais
decrescendo em tamanho e similares às brácteas inferiores.
Inflorescência 5-10 cm compr.; brácteas ovado-lanceoladas, 1.8-3.1 x 0.8-1 cm, caudadas, as inferiores aproximadamente do mesmo tamanho do ovário e pedicelo, decrescendo em tamanho em direção ao ápice da inflorescência, verdes. Flores (1)2-5(7), patentes, sobrepostas, grande para o gênero, verde-esbranquiçadas; ovário e pedicelo 2.5-3.5 cm compr., ligeiramente arqueado, verde; sépalas mucronadas, verde claro com a margem esbranquiçada, os feixes condutores discretamente destacados em verde mais escuro; sépala dorsal côncava, quando achatada elíptica, 12-16 x 8-13 mm; sépalas laterais obliquamente oblongas, 16-20 x 4.5-6.5 mm, acudas, deflexas nas flores completamente abertas; pétalas bipartidas, base branca ou creme, verde em direção ao ápice; segmento posterior linear-falcado, 13-15 x 2-3 mm, acudo, base livre, em direção ao ápice conivente com a sépala dorsal; segmento anterior linear-filiforme, 25-43 mm compr., 2-2.5(2.8) vezes mais longo do que o segmento posterior, projetado para frente e de certo modo paralelo aos segmentos laterais do labelo; labelo tripartido, projetado para frente, base branca ou creme e verde claro em direção ao ápice dos segmentos; base indivisa, 2-3 x 2-3 mm; segmentos laterais linear-filiformes, 32-51 mm compr., 1.7-2.3 vezes mais longos do que o segmento mediano; segmento mediano mais curto e mais largo, linear-ligulado, 17-24 x 1.5(2) mm, o ápice dobrado para cima; calcar pendente, clavado, livre das brácteas, ligeira a fortemente curvado para frente, mais estreito na base, progressivamente espessado para o ápice, 3.3-3.8 cm compr., base 1.5 mm larg., ápice 2.5--3 mm larg., verde; coluna 6-10 mm compr.; anteras ca. 4-6 mm compr., branco-amarelada; canais das anteras 7-9 mm compr, delicados, eretos, um tanto perpendiculares às anteras; conectivo emarginado, verde; lobo mediano do rostelo proeminente, alongado, formando uma estrutura em formato de túnel, projetado além das anteras, ca. 10-11 mm compr. incluindo os braços laterais, 6-7 mm de altura no ápice, carnoso, cuspidado, agudo, verde claro, a base branca, discretamente dobrada, formando um sulco discreto que se estende através dos braços laterais e que suporta os canais das anteras; braços do rostelo brancos, 3-4 mm compr.; aurículos eretos, ca. 2.5-3 mm de compr., carnosos, achatados, verrugosos em direção ao ápice, brancos; estigmas 2, paralelos, 6-8 mm compr. incluindo a base, a superfície receptiva 3-4 x 2-2.5 mm, projetada para frente, não involuta, superfície superior convexa, verde claro; polinários 2, amarelo, viscidio ca. 1 x 1 mm, espaçados 3-5 mm; caudículos alongados, 8-10 mm compr., ca. de 2-3 mm do ápice dobrados 90o; políneas 4, 3-4 x 1-1.3 mm, lateralmente achatadas, perpendiculares ao eixo central dos caudículos. Fruto não examinado. |
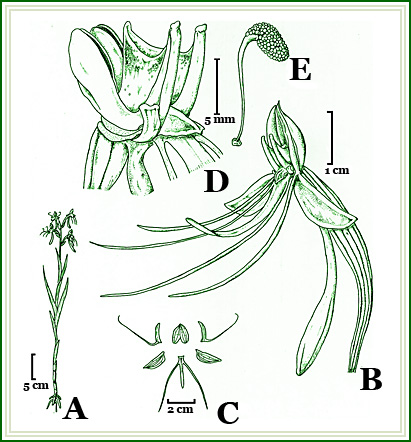 Figura 1. Habenaria pabstii Batista & Bianchetti. A. Habito, B. Flor - vista lateral, C. Perianto dissecado, D. Coluna - vista lateral, E. Polinário Desenhado do material tipo, Bianchetti et al. 1489, por Eduardo Gonçalves. |
Etimologia.
O nome da espécie é uma homenagem a Guido F. J. Pabst (1914-1980), o saudoso e conhecido orquidólogo brasileiro, autor do Orchidaceae Brasilienses, que contribuiu enormemente para o conhecimento das orquídeas do Brasil central e descreveu algumas espécies novas de Habenaria. Distribuição. Habenaria pabstii é uma espécie ocasional, conhecida até o momento apenas do Distrito Federal e do estado de Goiás. A espécie também deve ser esperada para o oeste de Minas Gerais, região floristicamente relacionada à região nuclear do bioma Cerrado e que compartilha muitas espécies em comum com o Distrito Federal e Goiás e provavelmente também no norte do estado do Tocantins, região próxima à área de distribuição conhecida da espécie. Habitat, Ecologia e Fenologia. Habenaria pabstii é encontrada mais comumente em campos limpos estacionalmente úmidos, em solos hidromórficos ou arenosos. Ocasionalmente, também pode ser encontrada sob condições mais secas ou, mais raramente, em solo permanentemente úmido, com água sobre a superfície. De modo similar a grande maioria das espécies de Habenaria do Brasil central, a floração ocorre durante o período chuvoso, de dezembro a março, estendendo-se do auge até quase o final do período chuvoso. Freqüentemente, as plantas aparecem em grande número, sendo que aparentemente apenas as mais velhas e mais desenvolvidas florescem. As flores geralmente não duram mais do que alguns poucos dias no campo. Adicionalmente, como várias outras espécies de Habenaria e orquídeas terrestres de campestres, a floração da espécie é muito favorecida por incêndios campestres (Jones, 1993; Oliveira et al. 1996). Grandes concentrações de plantas floridas são geralmente encontradas em áreas queimadas, embora ocasionalmente algumas plantas floridas também possam ser encontradas em áreas não queimadas. O mecanismo de polinização e o polinizador da espécie são desconhecidos. |
 |
O
primeiro a coletar H. pabstii foi, aparentemente, Ezechias P.
Heringer em 1961 no Distrito Federal. A análise de material seco
coletado naquela época e examinado por Guido F. J. Pabst revelou
que Pabst identificou erradamente a espécie como H. fastor
Warm., H. candolleana Cogn., H. urbaniana Cogn. e H.
vaupellii Reich.f. & Warm. Provavelmente estes enganos impediram
que H. pabstii fosse identificada antes como uma espécie
nova. Entre as Habenaria brasileiras, H. pabstii é similar na morfologia e tamanho das flores a um grupo de espécies na seção Macroceratitae Kraenzlin, representado por H. bractescens Lindl., H. fastor Warm., H. gourlieana Gill. ex Lindl., H. johanennsis Barb. Rodr., H. longicauda Hook. e H. macronectar (Vell.) Hoehne, dai as identificações anteriores de Pabst como H. fastor e H. vaupellii (um sinônimo de H. johannensis). Todavia, distinto de todas estas espécies, H. pabstii tem o calcar mais curto, livre, projetado para frente, e estigmas mais curtos, não involutos, que são paralelamente juntos, formando uma superfície receptiva única, voltada para cima e para frente. Em todas estas outras espécies o calcar é pendente, completa ou parcialmente envolvido pelas brácteas e mais longo do que o ovário e pedicelo, variando de 4 a mais de 15 cm, e atingindo até 25 cm em H. longicauda (Snuverink & Westra, 1983). Também, os estigmas são sempre mais longos (a parte livre varia entre 6-9 mm de comprimento em outras espécies com flor de tamanho similar a H. pabstii), separados e em contato apenas no ápice, as margens involutas, isto é, dobradas em espiral para dentro (Stearn, 1983) e a superfície receptiva no ápice voltada principalmente para os lados. Assim, embora H. pabstii indubitavelmente compartilhe algum grau de semelhança com estas espécies, difere em importantes detalhes da estrutura da coluna e a sua classificação infragenérica exata não é clara. Como a atual classificação infragenérica de Habenaria precisa de um rearranjo radical (Pridgeon et al. 2001), estudos futuros incluindo análises moleculares e genéticas possivelmente serão necessárias para esclarecer a relação entre H. pabstii, as espécies na seção Macroceratitae e outras espécies no gênero. Entre outras espécies na seção Macroceratitae, H. pabstii também é similar na morfologia geral das flores a H. quinqueseta (Michx.) A. Eaton. Todavia, esta espécie tem as folhas mais patentes, ovadas a lanceoladas e mais largas (2-6 cm de largura); pétalas e labelo brancos; o calcar mais longo e pendente (5-10 cm de comprimento, atingindo até 18 cm em H. quinqueseta var. macroceratitis (Willd.) Luer); e o lobo mediano do rostelo menor, completamente posicionado entre as anteras (Luer, 1972; Correll, 1978). Adicionalmente, H. quinqueseta tem uma distribuição geográfica distinta, ocorrendo no sul da América do Norte, América Central, Antilhas, e norte da América do Sul, nunca tendo sido registrada no Brasil. Além das espécies na seção Macroceratitae, a única outra espécie brasileira com a qual H. pabstii é similar e poderia ser confundida é H. urbaniana (seção Nudae Cogniaux). Nas duas espécies as flores são grandes e os segmentos laterais das pétalas e labelo longos e filiformes. Entretanto, H. urbaniana é vegetativamente mais alta (86-122 cm de altura), tem folhas lanceoladas com até 2 cm de largura; a inflorescência mais longa (12-19 cm) e mais multiflora ((6)12 a 25 flores); flores geralmente menores com o segmento anterior das pétalas proporcionalmente menos desenvolvido (segmento anterior 19-33 mm de comprimento, 1.6-1.9 vezes mais longo do que o segmento posterior); calcar mais curto (ca. 2-2.5 cm), que não ultrapassa o ovário e pedicelo em comprimento e proeminentemente espessado para ápice (1 mm de largura na base, 3-4 mm de largura no ápice). Adicionalmente, a estrutura da coluna entre as duas espécies é muito diferente, sendo o lobo mediano do rostelo da H. urbaniana menor (cerca de 1.5 mm de comprimento e 2 mm de altura), triangular e completamente disposto entre as anteras. Por ultimo, plantas de H. pabstii tornam-se negras quando secas ou fixadas em etanol, enquanto as flores de H. urbaniana e das espécies na seção Macroceratitae tornam-se marrons. A identificação de H. pabstii como H. candolleana é fortuita uma vez que esta espécie tem flores bem menores (sépala dorsal e segmentos laterais do labelo com 6-8 mm de comprimento) e coluna com estrutura muito distinta. Na estrutura da coluna, principalmente em relação à morfologia do labelo, H. pabstii é similar a H. johannensis. Nas duas espécies o lobo mediano do rostelo forma uma estrutura proeminente, em forma de túnel, que é parcialmente ereta e projetada além das anteras. Apesar desta similaridade, não se sabe, e seria interessante investigar, se as duas espécies também apresentam semelhanças nos seus mecanismos de polinização. |
 |
 |
Parátipos.
BRASIL. Agradecimentos.
|
|
- Batista,
J. A. N. & L. B. Bianchetti. 2003. Lista atualizada das Orchidaceae
do Distrito Federal, Brasil. Acta Bot. Bras. 17(2): 183-201. |
|