 |
Some considerations about Venezuelan unifoliate Cattleya by Jan Pahl, Gerardo Castiglione and Rafael Vaamonde |
Introduction
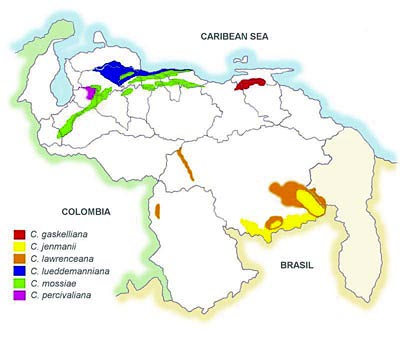
Venezuela
is a well-represented country inside genus Cattleya,
subgenus Cattleya, a group of orchids that
we often rename as unifoliate Cattleya.
Venezuela has 6 of the 20-21 recognized species,
5 endemic, and 1 almost completely endemic. Only
Colombia matches Venezuela in the number of large
species inside this subgenera.
Genetic
evidence show us that unifoliates, bifoliates Cattleyas and Brassavolas
are indeed closely-related groups, and that Cattleya derived
quite recently from a single Brassavola-like species that had
the chance to cross the prehistoric oceanic gap between Central
and South America. In which it found new ecological niches, evolving
into a kind of Proto-Cattleya, which was the common ancestor to
the two related Cattleya groups we have today.
As
we said, both Cattleya groups recently
evolved from a common ancestor, and species beneath
each group are even more recent in history, one
that, if taking into account present unifoliate
species, possibly started on the remaining archaic
unifoliate populations from past diseminations
that occurred during the global climatic changes
in the previous glacial and interglacial eras
in late Pleistocene.
Colonization
with posterior isolations are even occurring today, a feature
that is evident between the two mayor and not completely continuous
populations of Cattleya dowiana from Colombia/south-Panama
(Cattleya dowiana ssp aurea), and Costa Rica/north-Panama.
(Cattleya dowiana ssp dowiana), or between the two
isolated populations Cattleya labiata in Brazil. Those
quite recent diseminations, along with posterior ecological and
geographic isolation, are responsible for the richness of the
species we see today in wild populations of unifoliates found
even outside the northern Andean initial cradle.
Taking
this into consideration, there is no wonder in
why almost all large unifoliates share more similarities
than differences, genetically and anatomically
speaking, but there is no need to have clear stable
differences to assume speciation has actually
occurred, clear stable differences is a luxury
that only old, well-established genera or subgenera
have.
Venezuelan
Unifoliates, like the majority of their sisters,
prefer to inhabit mountain ranges from the low
land slopes and hills, up to altitudes of almost
2000 meters above sea level where they are exposed
to elements. We can find a species adapted to
almost any of the fore mentioned circumstances,
inhabiting almost every mayor mountainous range
from the Andean region, to the Coastal range,
and from the Costal range, to the Turimiquire
range, and from there, to the highlands of the
Guayana plateau. Only the Perijá range,
located in the western border with Colombia, lacks
her own indigenous species, an unsolved mystery
considering that Perijá has almost all
of the preferred conditions this subgenus exhibits.
|
| Cattleya lueddemanniana Rchb. f. 1854 |
 C. lueddemanniana ´María Gabiela´ x ´Maruja´ |
Cattleya lueddemanniana is one of the easiest unifoliates to recognize from similar species and possible primary hybrids, is also one of the best shaped Cattleya from Venezuela. The flower column is enclosed by the lip in a distinctive way that forms two noticeable side-lobes where ocular yellow spots are located on each side, with very few exceptions found on some rare cegatas. The flower’s lip is normally flat and rounded, with noticeable frilled edges. With the exception of Albas, they have amethyst-purple markings that run without interruption from the inner part of the tube, reaching the frontal spot of the labellum. They also have a particular flower-column with a bi-horned well-developed anther that is very easy to notice, especially when compared with the more "hook-like" anther of other unifoliates, only very few exceptions, many of them candidates for possible natural hybrids, have partially developed ones. |
|
Cattleya
lueddemanniana inhabits two different types
of habitat. The first one is the lowland xemi-xerophytic,
deciduous, and semi-deciduous, lowland forests
in the Caribbean facing slopes of the "Cordillera
de la Costa" range. Located in the northernmost
part of this range, between a confined band
of altitudes that range from 50/100 meters above
sea level, to 500 meters above sea level (maybe
more in some places), where the interaction
of the mountain range with the proximity of
the sea some humidity is provided even in the
dry season. In the western part of the "Cordillera
de la Costa", the species moves inland
to the lowland forests of the Yaracuy depression,
at the end of this natural depression, the species
reaches the second type of habitat, the foot
hills of "Los Andes" range, in an
area confined by Carora depression (west), Mirimire
valley (North) and Andes range (south), in this
place, C. lueddemanniana is present at
higher altitudes, between 400 and 700 meters
above sea level, in open xerophytic, xemi-xerophytic,
and deciduous forests created by the orographic
shadow of the Andes range, in a type of forests
manly composed by small trees, thorny bushes
and columnar cactuses.
Since
it inhabits two different types of habitat,
the species has two distinctive populations
with unique adaptations to endure different
ecological needs, and for that same reasons,
the species forms two different biotypes, with
a transition population located on the Yaracuy
depression. This transition population has intermediate
characteristics, and forms a natural cline between
both continuous populations. Since this transition
biotype is located in a very large and representative
area (central Yaracuy and north-west Carabobo
state), continuity between populations ensure
genetic flow, some collectors and growers do
not consider each distinctive population as
clearly separated subspecies.
The
coastal biotype forms large colonies in small to medium sized
trees, it somehow has more coriaceous leaves than the typical
Cattleya, due to that lueddemanniana colonies remain
at least partially unprotected from direct sunlight when trees
lose their leaves since in the cooler months of the dry season.
Plants remain well protected in the warmer rainy season, when
trees have leaves. |
Normally
coastal Cattleya lueddemanniana has big
flowers that can reach 16-18 centimeters, or
even more in exceptional cultivars, almost all
are light colored (suaves and suavissimas).
They also have a wide, normally very frilled
labellum with very light, almost washed yellow
ocular spots, more cream colored than real yellow.
The frontal spot of the lip is normally sprinkled,
many of them by hundreds of small amethyst-purple
points, or has rather small purple blotches
surrounded by sprinkled edges. Some plants are
known to have petals and sepals with very light
color, sometimes splashed (aquinii), with the
lip´s spot completely solid and velvety
up to the throat, with very intense yellow colored
ocular spots, sometimes masked by the purple
spot, these clones are known as “solid
lip”. Also, it’s well-known that
almost all of the albas, semialbas and all caeruleas
come from this area. Normally the best shaped
lueddemannianas come from this biotype; even
today some wild collected plants
could reach show standards.
|
 Cattleya lueddemanniana ´Oscura´ |
|
Xerophytic biotype lueddemanniana tolerate a longer dry season,
more heat, and less humidity, than coastal Cattleya lueddemanniana,
it forms large colonies in medium to small sized trees and thorny
bushes, even exposed to full sun on columnar cactuses. In Venezuela
this biotype is known as “Larense biotype” because
the main population occur inside Lara state.
The
xerophytic biotype has even more coriaceous leaves
than the coastal byotipe, more burgundy spots,
especially in new growths, and to endure dryer
and warmer conditions the overall vegetative aspects
of the plant tend to be more notorious in special
adaptations. Normally, xerophytic C. lueddemanniana
have medium to small-sized flowers (11 to 16 cm).
This biotype has narrow petals and erect "shoulders",
making the coastal biotype in terms of shape and
size far beyond better than the xerophytic biotype.
But this biotype is appreciated for other goods,
in terms of coloration the xerophytic biotype
has very intense colored flowers (roseas, purpureas
and rubras), this means xerophytic lueddemannianas
have a very desirable genetic stock that holds
the most intense colors of the unifoliate Cattleyas.
The labellum of this biotype tends to be rather
small and not so well shaped, normally the amethyst
purple blotch in the labellum is not solid, but
very densely striated or veined and very dark.
Colors are very intense to almost iridescent,
and the ocular yellow spots tend to be bright
colored (canary yellow).
The
flowering season begins in November and ends in
March, with a peak in February that can vary somehow
in cultivation due to cultural and climatic factors,
such as lack of resting period, or because of
cooler conditions. Given this, the cultivated
C. lueddemanniana can flower a second time in
the year, normally between August and September.
In the wild they remain dormant for several months
but in cultivation they don’t have a well-defined
rest period. Flowers develop from a green single
sheath, rarely without, or from old dry ones.
2 to 4 flowers grow from every spike. Exceptionally,
some cultivars can reach up to 5 flowers.
Is
difficult to write about how a C. lueddemanniana
should look in cultivation, we can only say that
the desired goal for many commercial breeders,
is to succeed in the best combination from each
mayor biotype, in other words, something that
could look exactly alike an enhanced flower from
the transition zone, with shape, size, and lip
pattern, similar to a coastal lueddemanniana,
and the bright deep color intensity of the xerophytic
C. lueddemanniana.
Some outstanding clones are: - coastal type: ´Maruja´, ´Haydé´, ´Calex´; - solid lip clones: ´Tibisay´, ´Francis´, ´Centellas´; - xerophytic type: ´Yamilé´, ´Cecilia´, ´Morella´, - alba: ´Morocha´, ´Candia´, - semialba: ´Mamacita´, ´Isabel Nataly´, ´Stanley´s´, ´Orquilara´; - caerulea: ´Mariauxi´, ´Siquisique´, ´Francisco´, ´Amparo´. |
 Cattleya lueddemanniana ´Maruja´ |
 Cattleya lueddemanniana ´Centellas´ |

Cattleya lueddemanniana alba ´Candelaria´ x (´Morocha´x ´Paola´) |
 Cattleya lueddemanniana semi-alba ´Isabel Nataly´ |

Cattleya lueddemanniana caerulea ´Mariauxi´ |
| Cattleya mossiae Hooker. 1838 |

Cattleya mossiae ´Carlo Aulisi´ x self |
C.
mossiae is an outstanding species which is easy
to recognize. Two features that make this species
unique are: A broad lip which is very crisped and
veined and broad petals which tend to fall. These
characteristics may be present in some degree in
wild plants as well as in cultivated plants. Despite
its flower shape (which is not the best of all the
Venezuelan cattleyas), the C. mossiae was
declared in 1951 Venezuela’s national flower.
The reasons that prevailed to select C. mossiae
over other Venezuelan cattleyas were: this specie
lives near some of the most populated areas in Venezuela;
its blooming season peak during the Easter Holidays
and Mother’s day; plenty of flowers are produced
that last up to four weeks. C. mossiae is endemic to Venezuela. It grows in a wide area along a mountain range called Cordillera de la Costa, either on the northern facing slopes or the southern facing slopes. |
|
It can also be found growing on all the south-facing
slopes of the Venezuelan Andes. It prefers to
grow in semi-deciduous to rain forest, from 800
to 1500 meters above sea level. The Cordillera
de la Costa is a mountain range which extends
from the eastern part of Miranda State (near Caracas)
to the western part of Yaracuy State. C. mossiae
reappears again near the Bordering States of Lara
and Portuguesa, and it extends along the Andes
Mountains up to Táchira State, near the
Colombian border, where the Táchira depression
establishes a natural barrier.
C. mossiae grows especially on large open trees like Bucares (Erythrina sp), Ingas, and Ceibas. It prefers to grow in the upper branches, forming large colonies, and protected from direct sunlight. It is also possible to find them growing on rocks, where the surrounding vegetation protects them from direct sunlight. C. mossiae normally grows near rivers or creeks which provide them with enough humidity to survive the dry seasons. |

Cattleya mossiae caerulea ´San Luis´ |
This
means that C. mossiae prefers the temperate conditions
which predominate in these mountain ranges. Despite the widespread area of distribution, C. mossiae does not have well differentiated ecotypes, biotypes or subspecies. Only slight differences can be found between plants inside the mossiae complex. One of these differences is the blooming season between the Andes and the Coastal population. Plants from the Andes Mountains start their blooming season in early February, peak in March, and end by mid May. Plants from the Coastal range start to bloom in late April, peak in late May and ends in early July. C. mossiae flowers have a broad color range and the way that colors are mixed in the lip are countless. These color variations could be masking differences between Andean and coastal populations, or even semi-deciduous from cloudy forest populations. |
|
After
the blooming season is over, the plant forms new
leads which are completely developed by early
August. Then in mid august, it rests 4 to 8 months.
In many cases, the resting period ends when second
lead grows from the previous one. Usually this
happens in November or December. From January
to May, growths from the previous year form buds
inside a single sheath, which could be either
dry or green. The first growth usually blooms
from a dry sheath, and the second growth from
a green one. On the average both growths will
bloom at the same time, but it is also possible
that they will bloom at different dates, up to
3 months in difference.
A
single spike can carry from 2 to 7 flowers. Usually
the flowers measure more than 16 cm. in diameter.
Some cultivars can easily reach up to 20 cm in
diameter. That’s why large plants in full
bloom of C. mossiae make a striking view
either in the wild or in cultivation.
|
 Cattleya
mossiae ´Chispa´
Cattleya
mossiae ´Chispa´ |
|
The
color range in flowers of C. mossiae normally
is limited to light colored tones (roseas, suaves
and suavissimas). Dark colored flowers of the
purpurea or rubra type have never been collected
in the wild. The trade mark of C mossiae’s
lipcolors are: reds, ochers and intense canary
yellows tones that intensify other colors in the
lip. The purple blotch in the lip could be solid,
striated, variegated or veined (striata, variegata
and venosa) except in the alba form where only
yellow is present There also exist alba, semialba,
concolor and caerulea varieties.
Some well-known clones are: color: ´Natalia´, ´J.H. Patterson´, ´Canoabo´, semialba: ´Aurora´, ´Blanca´, ´Featherston´, ´Julieta´, alba: ´Barroeta´, ´Crusizio´, caerulea: ´Macayra´, ´Ayala´, ´María T.´, ´Lizette´. C.
mossiae rarely interbreeds with C. percivaliana
in the Andean mountains, forming the natural
hybrid known as Catteya xperegrine. This
natural hybrid does not present the better visual
aspects of its parents, and consequently rarely
taken into consideration. Thus little is known
about it. C xperegrine occur in the rainy
forest of the Trujillo state and the Lara-Trujillo
border, where both parental species overlap.
|

Cattleya mossiae semialba ´Patrizia´ |
In
some places in Lara and Yaracuy States, wild populations
of C. mossiae and C. lueddemanniana overlap
forming the natural hybrid C. xgravesiana.
This natural hybrid seems to be more common in
the wild than C. xperegrine. There are
no large introgression populations like the ones
that we can find between species like Guarianthe
skinneri and G. aurantiaca (Guarianthe
xguatemalensis). Only small introgressive
populations between C. mossiae and C.
lueddemanniana have been found in very few
places Also, there are few wild-collected plants
of C. lueddemanniana or C. mossiae in
cultivation that are examples of mistaken identities,
for example, C. lueddemanniana s/a “Caracas”
is a true C. xgravesiana collected from
the wild.
Large
populations of C. mossiae still remain in the
wild, especially in some parts of the Andes Mountains.
But deforestation and predation of this species,
even inside protected areas, puts this species
in great risk of
|
extinction.
In a few decades Cattleya mossiae could be
as scarce as C. lueddemanniana and C.
gaskelliana are today.
|



| Cattleya percivaliana O´Brien. 1883 |
Cattleya
percivaliana is a lovely species endemic
to the Venezuelan Andes located in the Trujillo
state. Unlike other labiate Cattleyas, C.
percivaliana prefers to grow on rocky cliffs
or small trees from 1400 - 2000 meters. Morning
mist and specially creeks near the cliffs, provides
the moisture needed by this species to grow
exposed to direct sunlight and strong winds.
C. percivaliana often grows associated with vegetation (ferns, mosses, likens, grass, small shrubs), so they never suffer from dehydration. C. percivaliana is also adapted to grow on rainy forests, many healthy colonies occur on this type of habitat. C. percivaliana is not distributed over a wide area, if we consider the orography of the Andes, the average height of land is quite higher than the maximum height tolerated by this species, so population continuity is not possible and the species is isolated at the northernmost part of the Venezuelan Andes, near the Carora depression. |
 Cattleya
percivaliana ´Morena´
Cattleya
percivaliana ´Morena´ |
|
Some
authors consider that C. percivaliana grows on
the south facing hills of the Venezuelan Andes,
located in the states of Mérida and Táchira,
but this is not true. On these hills only grows
C. mossiae. There are few reports claiming
that C. percivaliana also grows in the
border of Norte de Santander state, Colombia,
but since there is a natural bond between Venezuela’s
Andean people and Colombia, it is not uncommon
to see C. mendelii and C. percivaliana
at both sides of the border. C. percivaliana is the smallest of all large unifoliate, only in terms of flower size. Flowers in wild plants normally measure 11 to 12 cm. There is a variety known as grandiflora (large sized percivaliana), not common, that has become a standard for cultivated plants. Flowers in C. percivaliana var. grandiflora can measure 15-18 cm. The plant size of this species is similar to other labiate Cattleya type. Usually a spike can carry 2 to 4 flowers. In late February, while other Venezuelan cattleyas are preparing to bloom, C. percivaliana awakes from a five month rest period. The new stems are ready to bloom from a green sheath in late August, but in many cases the buds can be formed any time from August through November. In the Venezuelan Andes, it is not unusual to see cultivated plants blooming all year round. C. percivaliana only blooms once per year but many cultural factors can change the normal blooming season. There is not a second growth like in C. mossiae. |

Cattleya percivaliana ´Enrique Jr.´ x ´Summit´ |
Two
characteristic makes C. percivaliana unique
within the genus, and easy to be recognized while
in bloom. The first one, and maybe the most significant,
is their scent like a recently cut grass that can
be considered as not pleasant, and also, the flower
shape that many plants found in the wild have, it
can be considered as almost perfect for Orchid Judgment.
The well-shaped flowers are very round, well presented,
and symmetrical. Petals in many cases present overlapping,
and good substance. The lip is small, frilled and
has a solid purple blotch that could vary in size
and shape. There is no evidence of striatas and
venosas within the species, only very few aquini
and variegate clons have been found. C. percivaliana
has a lip with amazing mixed colors that no other
large Cattleya can produce. There is a wide variety of art-shade colors present in the lip, between the most beautiful we can mention brown chocolate, guava, orange, red |
|
apple and Spanish gold among others. The type variety
has a purple color that can vary from medium to
very dark (variegate clons); alba, albescens, semialba
and caerulea clons are also known, but true concolor
or delicata forms exist. Some famous clons are: - type: ´Summit´, ´Karen´, ´Primavera´, ´Albert´s´, - semialba: ´Carache´, ´Sonia de Urbano´, - alba: ´Oro Cochano´, ´La Puerta´, ´Quintero´, - caerulea: ´Ondine´. We want to point out that the semialba plants known as ¨Farah Diva´, ´Jewel´ and ´Carache´ come from an unique and hudge plant property of the late Mr. Rafaelito Cañizales who lived in Carache, a small town in Trujillo state, Venezuela. |
|


| Cattleya gaskelliana Rchb. f. 1883 |
 Cattleya gaskelliana caerulea ´Divina´ |
One
of least widespread of all the Venezuelan Cattleyas
is the Cattleya gaskelliana. It thrives
only in a small area in the northeast of Venezuela,
called the Turimiquire range. This small mountain
range is located at the confluence of 3 bordering
States (Monagas, Sucre and Anzoátegui).
Like the C. mossiae, C. gaskelliana grows best
in altitudes that range from 800 to 1500 meters
above sea level. It grows in semideciduous to
ever green temperate forests, on medium to large
sized trees. In the Turimiquire range, rain
or morning mist is present all year round. Large populations of gaskelliana’s occur in the cloudy forests near the border of Monagas and Sucre States. Another population thrives in the northeastern border of Anzoátegui State, in a small area called Nuevo Mundo. Nuevo Mundo has a semideciduous forest, where C. gaskelliana can be found growing either on trees or rocks, forming large colonies. |
Some
authors consider that the population of C.
gaskelliana from Nuevo Mundo is a different
ecotype than those found in Monagas and Sucre
State. The Nuevo Mundo ecotype is characterized
by small sized pseudobulbs, and darker flower
colors. Some authors believe that this species can be found growing in Araya and Paria peninsulas, also in the Islands of Margarita and Trinidad. But many explorations have proven that no suitable habitats exist in these areas. Some reports from these areas are usually mistaken, as the possible sightings of this species always occurred in abandoned gardens and, in many cases, they were not even gaskelliana’s. |
|
C. gaskelliana has erect and broad leaves,
and normally produces new growths in late December.
These new growths are ready to bloom from April
through July, without a resting period. The
buds always form inside a very broad single
green sheath. Plants in cultivation can bloom
twice a year although a second flowering season
is not well defined. The Cattleya gaskelliana’s
flowers usually lack substance; they don’t
have upright petals, and are usually narrow.
Generally, it has a thick column which displays
two lateral tops that stand out from the anther.
On the average, flower colors can range from
suavissimas to suaves. The lip´s purple
blotch is quite variable, specially in color
intensity and size, usually triangular and almost
ever mixes with the yellow color and the brown-orange
veinings that come from the bottom of the throat.
The frontal part of the tube normally shows
cream or white tones, but since light flowers
are usual in this species, this feature is not
always quite evident. It´s relatively
easy to find alba, semialba, concolor and caerulea
varieties, but it´s difficult to find
dark colored flowers. |
 Cattleya gaskelliana semialba ´Calex´ AM-AOS |
| Some
famous clones are: - color: ´Elena´, ´Black Tiger´, ´Carmen´, - semialba: ´Calex´, ´Nuria´, ´Red Flame´, - alba: ´White Heritage´, ´Schnee´, ´Sara´, - caerulea: ´Blue Dragon´, ´Drago´, ´Aida´, ´Mimí´. |
| Cattleya jenmanii Rolfe 1906 |
Cattleya
jenmanii was described originally by Rolfe
in the 1906 Kew Bulletin, never again seen until
G.C.K. Dunsterville rediscovered it in 1969
from some plants that came from the southeastern
portion of Bolívar State. This beautiful
Cattleya shares her habitat with C.
lawrenceana in the Guayana plateau, Venezuela.
This species can be found growing near the Venezuelan-Brazilian-Guyanan
border (Roraima plateau), between upper Caura
river (state of Bolívar, Venezuela) and
Roraima (at the Venezuelan-Guyanan borderline).
Large populations of C. jenmanii occur
following the Venezuelan-Guyanan borderline,
between the 5º and 6º paralells. This
species is endemic to Venezuela, however, if
we consider that the Guyana plateau is a very
big unexplored area, there is a chance that
populations of this species could be found in
Brazil (there are few unconfirmed sightings),
or in Guyana, always near the Venezuelan border.
However C. jenmanii has not been reported
growing outside Venezuela. This species grows in a humid temperate area known as La Gran Sabana, with altitudes that range from 800 to 1200 meters above sea level. In this area, grasslands are predominant so, C. jenmanii only grows in the few forests that have successfully survived despite the low Ph and sandy soil conditions. It can also be found growing exposed to direct sunlight, in crevices or in the rocky cliffs called Tepuys (Tablelands). For some reasons introgressive populations between C. jenmanii and Cattleya lawrenceana have not been reported although populations of both species overlap in nature. We know of two collected plants of this natural hybrid whose name has not been registered yet. Two have been proposed: C. xgransabanensis and C. xcanaima. The shape of the flower is similar to an average C. jenmanii but with a lip very similar to a big C. lawrenceana. |
 Cattleya jenmanii ´Marie-Anne´
Cattleya jenmanii ´Marie-Anne´ |
The
leaves of C. jenmanii are erect and narrow,
with an elliptical shape that reminds us the
leaves of C. mossiae, although they sometimes
present a purple tone in the back; and, they
always present a single sheath, a characteristic
that gives us a first clue to help us differentiate
her from Cattleya labiata. |
 Cattleya jenmanii semialba
´A.M.C.´ x self
Cattleya jenmanii semialba
´A.M.C.´ x self |
|
The
flowers have a very sweet scent, with 12 to
17 cm in diameter, erect and elegant, but generally
with petals and sepals quite thin. The pattern
of color of the lip is very similar to Cattleya
labiata, being sometimes very difficult
to differentiate them. The throat is yellow
with brown-orange veinings towards the bottom,
the borders are delicately waved, rosy-purple
with the characteristics white “eyes”
on each side. The purple blotch can be just
lineata, estriata or completely solid. Like
in C. gaskelliana, the buds start to
grow inside the sheath before the new growth
is completely formed.
C. jenmanii has a predominant blomming season from February to April, with a second period between September and October. It is a relatively new species in cultivation and few quality clones are known. The color variety has a rose-purple coloring that can vary from quite clear (suaves) to very dark (rubras); alba, semialba, concolor, delicatas and caerulea clones are also known.
|


| Cattleya lawrenceana Rchb. f. 1885 |

Cattleya lawrenceana concolor ´Luz de Luna´ |
One
of the most atypical unifoliates of the Cattleya
genera is the C. lawrenceana, the pseudobulbs
are among de biggest of the genera. Plants usually
have pseudobulbs with a very distinctive redish
pigmentation, leaves also display a profusion
of burgundy and maroon spots specially those
that are subject to direct sunlight. Flowers
are easy to recognize as they are medium sized,
have a tubular labelum with a disc shaped fringeless
lip. Petals and sepals are usually narrow, flowers
have a greater resemblance to large Sophronitis
like S. purputata, tenebrosa, grandis
or lobata than to the other unifoliate cattleyas.
In
the Guayana Plateau the Cattleya lawrenceana
has a very wide and uneven area of distribution,
specially inside Bolívar State as populations
always follows mayor rivers and table mountains
formations called Tepuys wich are distant from
each other. One main population is located in
the Amazonas state (Sipapo Hill), and the others
are located unevenly on Roraima, Auyán-tepuy,
Kukenán, Chimata tepuys; High Caroní,
Carrao, Aponwao and Cuchivero rivers, also in
Kamara-ta Valley and Kanavayén area in
Bolívar State. The species is also found
in large quantities at the Guyana side of Roraima.
It also has been reported growing on the Brazilian
side, although confirmed only by few sightings.
|
|
Since
this species is quite adaptable and has a very
wide distribution, chances are that Cattleya
lawrenceana has more probabilities than
the Cattleya jenmanii to be found growing
in other regions than the known locations in
Venezuela, Brazil, and Guyana. Cattleya lawrenceana grows from lowlands (400 meters above sea level) up to altitudes of 1900 meters above sea level. In it´s natural habitats, it is a quite adaptable orchid growing on medium to large-sized trees in humid shadowy forest ranging from warm to temperate and cool humid forests. At high altitudes colonies adapt themselves to more open spaces, growing on cliffs exposed to full sunlight, with the exception of some protection found from surrounding vegetation. This particular area is very humid all year round, so C. lawrenceana (like C. jenmanii), even if they have to overcome the “dry season”, they always have the benefits of high humidity, mist and occasional showers, but since it is found growing on well vetilated spots, it suggests that they don’t stay wet for long periods of time. In cultivation C. lawrenceana is recognized to be difficult plant to grow. In the wild C. lawrenceana flowering season is from February to April. Like in many Cattleyas flowers arise from a pseudobulb that has rested for several months, 2 to 8 medium-sized rich rose-magenta flowers is the most common. Some cultivars might have even more flowers per spike (up to 12 in known multifloras), but that’s quite rare.
|

 |
| Authors
and plants' owners: Jan Pahl, orchidologist Gerardo Castiglione is judge in orchid shows and founder member of the Mérida Orchid Society. Rafael Vaamonde is member of the Venezuelan orchidology association, and responsible for the English version. The awarded plants showed here are from Gerardo Castiglione and Armando Mantellini private collection's. Armando Mantellini is one of the few Venezuelan pioneer growers responsible for the genetic melioration of the Venezuelan species and the Gerardo Castiglione's Cattleya collection is very well-know among Venezuelan orchidologists. |
|
Any
kind of reproduction (print, digital or anyone)
of any type of material of this site: texts,
layout, photos, images and others - is
strictly forbidden without previous written permission of the authors. Any solicitation or information by the e-mail: bo@sergioaraujo.com |