Possui graduação em História Natural pela Universidade Santa
Úrsula (1973), Mestrado em Ciências Biológicas (Botânica)
pelo Instituto Nacional de Pesquisas da Amazônia (1976) e Doutorado em Ciências
Biológicas (Botânica) pelo Instituto Nacional de Pesquisas da Amazônia
(1982). Atualmente é professor Titular aposentado da Universidade Federal
do Amazonas.
Tem experiência na área de Botânica, com ênfase em Taxonomia
- especialista na Família Orchidaceae, Biologia Floral, Fitogeografia -
Fitossociologia, Conservação Ambiental, Estudos de Impactos Ambientais,
Recuperação de áreas degradadas (www.orquideasdaamazonia.com.br)
ECOLOGIA
A família das orquídeas teve o seu maior desenvolvimento nas áreas
tropicais e a maior parte das espécies adaptou-se ao hábito epífito.
As relações delas com hospedeiros e as adaptações
para que pudessem viver como epífitas neles são muitas.
FORÓFITOS
São considerados forófitos aquelas árvores ricas em epífitas,
que possuem: estrutura da casca grossa e fendida; retenção de
umidade; nutrientes disponíveis; acumulam humos; não possuem produção
de substância alelopáticas (que inibem a germinação
de outras sementes) e tipo de arquitetura (disposição dos galhos
e penetração de luz).
Na
Amazônia brasileira, temos como exemplo a Bertholletia excelsa,
a Castanheira, a Hevea brasiliensis, a Seringueira, a Ceiba pentandra,
a Sumaumeira e a Aldina heterophyla, o Macucú, entre muitas
outras. Em um trabalho que participei com Jefferson da Cruz no Campo Petrolífero
do rio Urucu, Município de Coari, no médio Solimões,
de 86 árvores pertencentes a 63 espécies, 16 espécies
foram consideradas forófitos.
Nesses trabalhos, (CRUZ e BRAGA, 1976 e 1997) identificamos 29 espécies
de orquídeas.
Os estudos ecológicos existentes costumam relacionar a ocorrência
das orquídeas com o tipo de clima, vegetação, altitude,
altura no interior da floresta, e espécie de árvore hospedeira.
A existência de um gradiente microclimático vertical dentro da
floresta onde a intensidade luminosa, temperatura e circulação
atmosférica aumentam no sentido solo-dossel, é irrefutável.
Esse gradiente não é constante e varia de um local para outro,
devido à estratificação irregular das árvores e
às diferenças específicas e individuais na densidade de
suas copas. A interceptação de chuva também varia diretamente
com a densidade das copas, além de depender, como é óbvio,
do total de precipitação. Esses e outros fatores influenciam enormemente
no padrão de distribuição espacial dos indivíduos
de uma população.
A fim de que possamos entender a ocupação dos diversos habitas,
precisamos mencionar as várias adaptações e os fatores
que interagem:

1) As sementes das orquídeas são muito leves e são facilmente
transportáveis a grandes distâncias pelo vento e pela água.
Isso não só ajuda à dispersão das sementes, como
também permite que as plantas se estabeleçam em habitats bastante
específicos, em lugares precisos e árvores determinadas.
2) As sementes dessas plantas são muito sensíveis aos fatores
físicos e químicos. Entre os físicos citamos: umidade relativa
do ar, intensidade luminosa, arquitetura das árvores e tipos de substratos.
Entre os químicos destacamos: os nutrientes disponíveis, substâncias
alelopáticas e pH.
Dentro de uma determinada comunidade os gradientes microclimáticos são
bem evidentes. Podemos então dividir as plantas, quanto à sua
necessidade de 1uz, em: umbrófilas as que crescem no chão e nos
troncos das árvores; semi-heliófilas as que crescem margeando
a vegetação ou nos ga1hos; e heliófilas as que crescem
em pleno sol.
3) Uma das características dessa família é a ausência
de endosperma nas sementes e para que o desenvolvimento das plântulas
tenha pleno êxito, é necessária uma relação
de simbiose entre essas e os fungos do gênero Rhizoctonia.
4) Com o epifitismo, a obtenção de nutrientes, bem como o suprimento
de água tornaram-se críticos. A fim de que o balanço hídrico
fosse mantido, foram necessárias adaptações xerofíticas
como: velame, pseudobulbos, folhas carnosas e metabolismo ácido crassuláceo
(C.A.M.) em muitas dessas epífitas.
Para que o suprimento de minerais ocorresse, adaptações como o
grande número de raízes, rizomas e pseudobulbos permitiram que
material orgânico se acumulasse entre eles, ocorrendo assim à formação
do que se chama de húmus. Não podemos nos esquecer também
da grande importância das formigas do gênero Azteca e outros animais
que fazem os seus ninhos na base destas plantas, acumulando muita matéria
orgânica.
O suprimento de nitrogênio nessas plantas ocorre pelas chuvas ou pelas
gotas de água que pingam das folhas depois de uma chuva, em forma de
amônia. Essas gotas de água contêm três vezes a quantidade
de NH3 encontrado na água da chuva.
No quadro que se segue reunimos os fatores que afetam a distribuição
de orquídeas
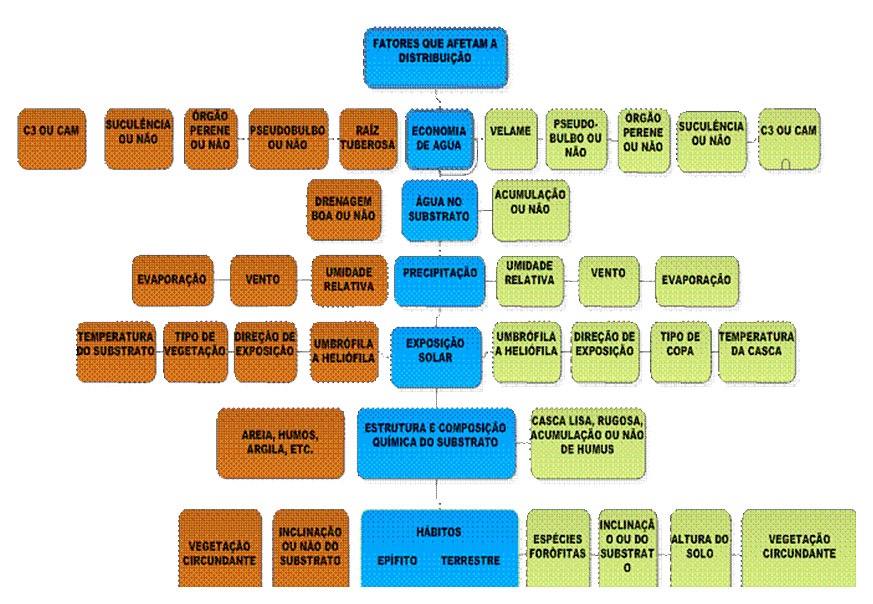
Por último abordamos a diferenciação e hibridação
em orquídeas da Amazônia.
A espécie em questão, Cattleya eldorado, é muito
variável e pode-se notar uma diferença marcante nos indivíduos
que ocorrem no Igapó ou Campinas próximas das margens dos rios,
em relação aqueles que ocorrem em Campinas da Terra-Firme.
Os primeiros possuem plantas mais robustas e flores maiores do que as dos últimos.
É importante citar que na época do descobrimento da espécie
e suas variedades a Terra Firme ainda não tinha sido desbravada por estradas,
o que a tornava de difícil acesso. Na Lindenia, inclusive é citado,
que as primeiras plantas que floriram provieram das margens do rio Negro e na
Martii Flora Brasiliensis, do rio Negro e Urubu.
O período da floração dessas plantas também são
diferentes.
No Igapó florescem de agosto a outubro e na Campina florescem geralmente
de dezembro a fevereiro.

Atualmente nas proximidades de Manaus, as Campinas foram destruídas
e as Campinas (que visitei) estavam em Terra-Firme entre 60 a 90 km do
perímetro da cidade. Pode ser este o mistério do porque
daquelas variedades citadas na referida obra só terem sido reencontradas
recentemente.
Dito isso, penso que temos dois ecótipos que estão em processo
de diferenciação ou mesmo podendo ser considerados como vicariantes
de duas subespécies.
Entretanto, ambas populações possuem o mesmo perfume forte característico.
Em relação ao formato das flores também é muito
variável e a maioria das flores não mostram boa armação.
Quanto a coloração variam do rosa - claro ao rosa - escuro ou
de flores brancas.
Todas as Cattleya eldorado Lindén possuem o labelo com a fauce maculada
de laranja. De modo geral, é possível encontrar formas intermediárias
entre as variedades o que sugere a hibridação destas em condições
naturais.
Como sabemos, a flora das Campinas Amazônicas foi em grande parte derivada
do Escudo das Guianas. Por outro lado, com a retração da Floresta
de Terra-Firme no Pleistoceno, a ligação entre os tipos não
florestais foi ampliada.
No Quaternário a Floresta tornou-se a expandir propiciando uma diferenciação
e isolamento relativos entre as tipologias vegetacionais, pois o isolamento
não é total como em ilhas oceânicas.
Assim existem ocorrências de híbridos como a Cattleya
X brymeriana Reichb.f. (Cattleya eldorado X Cattleya violacea)
X Brassocattleya rubyii Braga (híbrido intergenérico
entre a Cattleya eldorado e Brassavola martiana Lindl.
No caso do último híbrido ocorre a formação de frutos
com sementes férteis e viáveis o que possibilita a ocorrência
de introgressão genética possibilitando variação
e especiação.

Esperamos com esse artigo ter contribuído para ilustrar como as orquídeas
ocupam os nossos ecossistemas e as especificidades que possuem de habitat, o
que faz com que possa ficar vulnerável a extinção.
• Sugestões para leitura
BENZING, D. H. - 1986. The vegetatives basis of vascular epphytism. Selbyana,
9(1):23-43.
-------1987. Vascular epiphytism: taxonomic participation and adaptative diversity.
Ann. Missouri Bot. Gard., 74(2):183-204.
BONATES, L.C. de M. - 1987. Aspectos Ecofisiológicos de algumas Orchidaceae
que ocorrem no estrato terrestre da vegetação de Campina da Reserva
Biológica do INPA, Km 45. Tese de mestrado apresentada ao curso de pós-graduação
do INPA-UFAM. 269p.
BONATES, L.C. de M. & BRAGA, P.I.S. - 1992. Estudos Ecofisiológicos
de Orchidaceae da Amazônia I. - Identificação da via C3
e CAM em quatorze espécies que vegetam no estrato terrestre de uma campina
da Amazônia central. Bol. Mus. Para. Emílio Goeldi, sér.
Bot., 8(2): 163-189.
BRAGA, M.M.N. & BRAGA, P.I.S. – 1976. Estudos Ecológicos na
Campina da Reserva Biológica INPA-SUFRAMA, Km 45. Acta Amazon. Manaus,
5(3): 247-260.
BRAGA, P.I.S. - 1976. Atração de abelhas polinizadoras de Orchidaceae
com o auxílio de iscas-odores na Campina, Campinarana e Floresta Tropical
úmida da região de Manaus. Ciênc. Cult, 28(7): 767- 773.
------- - 1977. Aspectos Biológicos das Orchidaceae de uma Campina da
Amazônia Central. Acta Amazon. Manaus , 7(2), Suplemento: 89p.
------- - 1978. Estudos da flora orquidológica do Estado do Amazonas
III- Brassocattleya rubyi Braga (Orchidaceae) híbrido novo da flora amazônica.
Acta Amazon. Manaus, 8(3): 371-378.
------- - 1979. Subdivisão fitogeográfica, tipos de vegetação,
conservação e inventário florístico da floresta
amazônica. In: Estratégias para a política florestal na
Amazônia brasileira. Acta Amazon. Manaus, (9(4): 53-80. Suplemento.
------- -1981a. Orquídeas da Amazônia brasileira. Bol. Assoc. Orquid.
Amazon., 1(2): 16-24.
------- -1981b. Orquídeas das Campinas da Amazônia brasileira.
Bradea, 3(23): 170-173.
------- - 1981c. Orquídeas das Campinas da Amazônia brasileira.
In: Anais do I Encontro de Orquidófilos e Orquidólogos do Brasil,
Rio de Janeiro, Expressão e Cultura. p. 19-43.
------- - 1982a. - Orquídeas da Amazônia brasileira II. Bol. Assoc.
Orquid. Amazon., 2(1): 85-88.
------- - 1982b. Aspectos biológicos das Orchidaceae de uma Campina da
Amazônia Central. II- Fitogeografia das Campinas da Amazônia brasileira.
Tese de doutorado apresentada no curso de pós-graduação
do INPA-UFAM. 345 p.
------- - 1983. Fitossociologia da Família Orchidaceae I. Bol. Assoc.
Orquid. Amazon., 3(5): 185-188.
------- - 1987a. Orquídeas- Entrada e Dispersão na Amazônia.
Ciênc. Hoje, 5(28): 44-51.
------- - 1987b. Orquídeas- Biologia Floral. Ciênc. Hoje, 5(28):
52-56.
------- -1994. Orquídeas da Amazônia Brasileira III. Bradea, 6(35):
293-296.
BRAGA, P.I.S. & VILHENA, R. - 1981. Estudos sobre a vegetação
de campinas amazônicas VII.- Anatomia ecológica de Epidendrum huebneri
Schltr. e Phthirusa micrantha Eichl. In: Anais do I Encontro de Orquidófilos
e Orquidólogos do Brasil. Rio de Janeiro, Expressão e Cultura.
p. 86-106.
BRAUN BLANQUET J.- 1979. Fitossociologia: bases para el estudio de las comunidades
vegetales. Madri, H. Blume Ediciones.
BRIEGER F.B., MAATSCH R. e SENGHASK. - 1971. Die pflantzem geographische verbreitung
der Orchideen, R. Schlechter (org.), Die Orchideen. Berlim, Paul Parey.
CAIN S.A.- 1944. Foundations of plant geography. Nova Iorque, Harper & Brothers.
CRUZ, J. da & BRAGA, P.I.S. - 1996. Considerações sobre a
relação das Orchidaceae Epifíticas com seus forófitos
em floresta de terra firme no campo petrolífero do rio Urucu, Médio
Solimões, Amazonas - Brasil. Rev. UA, Série: Ciências Biológicas,
Manaus, 1(1): 17-29.
--------- 1997. Aspectos taxonômicos e ecológicos de Orchidaceae
epífitas no Campo Petrolífero do Urucu, Amazonas - Brasil. Rev.
UA, Série: Ciências Biológicas, Manaus, 1(1): 1-129.
DUCKE, A. & BLACK, G. A. - 1954. Notas sobre a fitogeografia da Amazônia
brasileira. Bol. Técn. IAN, Belém, 29:1-62.
GOOD R.- 1974. - The geography of the flowering plants. Londres, Longman Group
Ltd.,
HOEHNE, F. C. - 1949. Iconografia de Orchidaceae do Brasil:1-301, 300 tab.
MADSON, M. -1977. Vascular epiphytes: Their systematic occurrence and salient
features. Selbyana, 5(2): 207-213.
MIRANDA, F. L. - 1982. As variedades da Cattleya eldorado Linden. Bol. Assoc.
Orquid. Amazon., 2(2): 91-95.
NORTHEN, R. T. - 1984. Their innocent past, their promising yet previlous future.
IN: Orchid Biology Reviews Perpectives. III. Edited by Joseph Arditti. Comstock
Publishing Associates. p. 19-26.
PABST, G.F.J. & DUNGS - 1975. Orchidaceae brasilienses, 1. Hildesheim, Brücke-Verlag
Kurt Schmersov. 408 p.
PABST, G.F.J. & DUNGS - 1977. Orchidaceae brasilienses, 2. Hildesheim, Brücke-Verlag
Kurt Schmersov. 418 p.
PRANCE G.T. - 1977.The phytogeographic subdivisions of Amazonia and their influence
on the selection of biological reserves", Extinction isforever. Nova Iorque,
New York Botanical Garden.
PRANCE ET ALII - 1979. Árvores de Manaus, Manaus, INPA. 312 p.
WAECHTER, J. L. - 1980. Estudo fitossociológico das orquídeas
epifíticas da Floresta paludosa do Faxinal, Torres, Rio Grande do Sul.
Porto Alegre, UFRGS, 104p. (diss. mestrado).
WENT, F. W. - 1940. Soziologie der Ephyten eines tropischen Urwaldes. An Jard.
Bot. Beutenzorg, 50:98 apud RICHARDS, R. E. - 1966. The Tropical rain forest,
an Ecological Study. Cambridge. p. 1-450.
WITHNER, C. L. - 1959. The orchids. A scientific survey, Ronald Press Company,
New York: 1-648.
|

