|
| Palavras-chave: Dungsia, Hadrolaelia, Hoffmannseggella, Laelia, Microlaelia , Brasil, Taxonomia. |
| O
gênero Laelia foi divido em várias entidades por diversos
autores, a partir das observações morfológicas. Um
estudo recente, baseado na análise molecular de um fragmento de ADN
ribossômico, confirma a existência de dois grupos bem distintos:
as espécies mexicanas (as verdadeiras espécies de Laelia)
e as espécies brasileiras que devem ser separadas das primeiras.
Uma importante congruência entre caracteres morfológicos, ecológicos e moleculares nos levam a restabelecer o gênero Hoffmannseggella H.G. Jones para nele colocar as espécies rupícolas, a reconhecer o caráter único da Laelia lundi Rchb. f. e a criar dois outros gêneros para as espécies restantes. Os notogêneros apropriados são criados e as novas combinações necessárias são publicadas. |
| Quando
o gênero Laelia foi criado, em l831, J. Lindley o descreveu
como próximo do gênero Cattleya, mas possuindo oito
polínias e não quatro e baseou-se em duas espécies
mexicanas anteriormente descritas como pertencente ao gênero Bletia
por La Llave & Lexarza, Bletia grandiglora e Bletia autumnalis,
deixando em separado, dentro do gênero Cattleya, a espécie
brasileira hoje conhecida como Laelia crispa. As Laelias são plantas epífitas ou litófitas, originárias do México (e norte da América Central) e do Brasil. Seus pseudobulbos são espessos, mais ou menos ocos (em algumas espécies mexicanas), claviformes, fusiformes, ovóides ou cilíndricos e possuem no ápice, uma ou duas (às vezes até mais) folhas geralmente coriáceas. A inflorescência é terminal e carrega, às vezes, uma única flor, às vezes diversas flores dispostas em racemo ou, mais raramente, em panícula. Com exceção das espécies rupícolas, as flores são, em geral, grandes e espetaculares, de cor bastante variada. As sépalas e pétalas são livres, bem abertas, explanadas ou onduladas. O labelo é livre ou apenas adnato à base da coluna, pode ser inteiro ou trilobado, formando um tubo em volta da coluna junto à base para, em seguida, se explanar, o disco do labelo é liso ou em lâminas. A coluna é alongada e apresenta um dente no ápice. A antera apiculada protege oito polínias cerosas, ovóides ou comprimidas, dispostas em 4 por célula. Pouco depois de haver criado o gênero, Lindley propôs uma primeira subdivisão e reconheceu particularmente uma seção Parvilflorae, brasileira (Lindley, l842). Desde então, diversos autores (Cogniaux, l901; Schlechter, l917; Pabst & Dungs, l975) destacaram o caráter heterogêneo do gênero Laelia senso lato, quer dizer, reagrupando as espécies mexicanas e as espécies brasileiras. Brieger (l981) deu um passo adiante e dividiu o gênero em dois subgêneros: o subgênero Eulaelia, que agrupa as espécies mexicanas, as "verdadeiras" Laelias já que a espécie-tipo (1) do gênero, Laelia speciosa (Kunth) Schlecther, faz parte dele; e o subgênero Brasiliensis, que agrupa as espécies brasileiras. Efetivamente, desde muito tempo, os orquidólogos assim como orquidófilos consideram que as espécies mexicanas de Laelia estão muito próximas de Schomburgkia Lindley e de Encyclia Hook do que as espécies brasileiras de Laelia. Por outro lado, no Brasil, considera-se que a espécie mais representativa do grupo brasileiro é a Laelia purpurata Lindley que, sob o ponto de vista vegetativo, se assemelha mais a uma Cattleya Lindley do que a uma Laelia mexicana. Mesmo entre as espécies brasileiras, as diferenças morfológicas são grandes e grupos mais homogêneos foram delimitados. Assim, H. G Jones (1968) criou o gênero Hoffmannseggella para as espécies rupícolas, mas esta concepção não foi seguida pelos taxonomistas. Prosseguindo o trabalho de classificação do gênero, Withner propôs, há pouco tempo, a criação de 3 subgêneros para organizar as espécies brasileiras de Laelia (Withner, l990); um destes subgêneros coincide, bem entendido, com o gênero Hoffmmannseggella H.G. Jones. Mas, talvez para evitar um tumulto no meio horticultural, Withner nunca chegou a colocar as espécies brasileiras neste gênero distinto. Mais recentemente, Halbinger e Soto apresentaram um estudo cladístico das relações entre os membros da chamada aliança Laelia-Cattleya com base em 64 caracteres morfológicos (Halbinger & Soto, l997). Esta análise confirma o distanciamento das espécies mexicanas das espécies brasileiras, a existência de um grupo natural " Laelia subgênero Parviflorae" (com um bootstrap - índice de confiabilidade - de 97%), mas falta resultado no que se refere aos grupos "Laelia subgênero Crispae" e "Sophronitis ". Todo este trabalho taxonômico, inicialmente baseado apenas nos caracteres morfológico das plantas, leva em conta, hoje em dia, outros caracteres, tanto intrínsecos (odor, época de floração) como extrínsecos (polinizadores, biogeografia...). A recente evolução das técnicas de análise molecular, permitindo acesso ao seqüenciamento de partes do ADN, multiplicou consideravelmente o número de caracteres disponíveis. Recentemente, van den Berg e seus co-autores estudaram a filogenia da subtribo Laeliinae através da análise molecular da seqüência ITS do ADN ribossômico (van den Berg et al. 2000). Este estudo confirma a existência, dentro do gênero Laelia sensu lato, de dois conjuntos bastante distantes no plano filogenético. Dados morfológicos, ecológicos e moleculares estão assim em concordância e esta congruência reforça a necessidade de uma separação das espécies brasileiras do gênero Laelia. van den Berg et al. foram, assim, os primeiros a dar este passo. A árvore filogenética relativa às espécies brasileiras de Laelia mostra claramente a existência de muitos clades bem delimitados (Van den Berg et al. op. cit. fig 6, página 109), como pode se ver com mais evidência na figura 1, reproduzindo esta árvore de maneira condensada. Os resultados levaram Van den Berg e seus co-autores a reagrupar dentro do gênero Sophronitis todas as espécies brasileiras de Laelia. Esta concepção, entretanto, leva, por um lado, a juntar dentro de uma mesma entidade, plantas morfologicamente muito diferentes e, por outro lado, a ignorar os trabalhos dos predecessores. Parece-nos necessário discuti-la e propor uma organização menos distanciada da concepção clássica. (1) O nome Bletia grandiflora La Llave & Lex. l825 sobre o qual foi baseada a espécie-tipo Laelia grandiflora (La Llave & Lex) Lindley, é, na verdade, um sinônimo de Bletia speciosa Kinth, anteriormente poublicado (l815). Este último epíteto tem, então, a prioridade. |
|
||
|
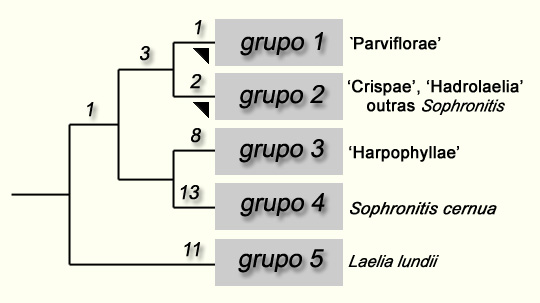
Figura
1: Cladograma das Laelias brasileiras
(adaptado de van der Berg et al., 2000) |
||
| O número colocado acima de cada ramificação (largura de Fitch) caracteriza o número de mudanças observadas na seqüência estudada ao longo desta ramificação. | ||
| |
| Antes de ir mais longe na discussão dos resultados apresentados por estes autores, é preciso relembrar algumas observações de ordem geral sobre a própria natureza do instrumento "filogenia molecular", observações que devem nos levar a manter a maior prudência na discussão dos resultados. |
| o estudo das relações filogenéticas baseadas num gene particular analisado dentro de um conjunto de espécies nos informa, com o máximo rigor, sobre a filogenia deste gene e não sobre a filogenia das espécies envolvidas; este problema foi e continua sendo avidamente debatido (ver principalmente a apresentação bem completa que fizeram Brower et al. l996); somente após ter obtido resultados concordantes sobre diversos genes, pode-se afirmar, sem grande perigo de erro, que a árvore filogenética assim construída se aplica também às espécies estudadas; |
| os caracteres moleculares do gene estudado não fornecem um resultado absoluto e incontestável (e, deste ponto de vista, a seqüência ITS, geralmente utilizada por razões de comodidade experimental, é particularmente frágil); ainda que, sem dúvida, menos subjetivo do que um estudo baseado na seleção de caracteres morfológicos, o método apresenta seus próprios limites e não dispensa o estudo atento de seus caracteres; ao contrário, ele deve servir de guia para retomar a análise morfológica dentro de uma ótica renovada, a fim de resolver eventuais conflitos entre os dados moleculares e os dados morfológicos (ver, por exemplo, a discussão de Lecointre & Le Guyader, 2001); |
| a seqüência analisada apresenta um nível baixo de variação de uma espécie para outra, principalmente no interior de cada um dos grupos esquematizados na figura 1; a própria confissão dos autores, <<os fracos níveis de divergência poderiam implicar num erro de amostragem como causa de colocações inesperadas>>. De toda maneira, esta característica traz uma fraca resolução na extremidade das ramificações e não permite o posicionamento apurado das espécies em relação às outras no interior de um grupo: notemos, por outro lado, que as variações registradas entre dois representantes da mesma espécie (que é o caso de, por exemplo, da Cattleya maxima, da Laelia xanthina ou da Sophronitis cernua) são, às vezes, mais importantes do que aquelas que separam espécies distintas. |
| observam-se,
num pequeno número de ramificações, freqüências
de mudança de estados muito elevadas (desenvolvimento em paralelo)
como na Cattleya maxima, Laelia fidelensis, Laelia virens,
Laelia perrinii e Sophronitis cernua; observou-se (é
o fenômeno conhecido pelo nome de atração das longas
ramificações) que, quando as velocidades de evolução
entre linhagens são muito diferentes umas das outras, as espécies
que evoluem mais rápido têm mais oportunidade de compartilhar
os estados de caractere por homoplasia do que por ascendência; em
conseqüência, as espécies que têm longas ramificações
(quer dizer, um grande número de variações), podem
ser reagrupadas dentro da árvore filogenética independentemente
de seu parentesco. (2) Notemos que os índices globais de homoplasia para a sub-tribo, no seu conjunto, são relativamente importantes, com um índice de coerência de 0,26 e um índice de retenção de 0,71. |
| certos clades são fragilmente sustentados pelos testes de coerência e de robusteza (porcentagem de bootstrap pouco superiores e, freqüentemente, inferiores a 50%); ainda aí, e os próprios autores assinalam que, nestas condições, é prudente, aguardar os resultados complementares. |
|
O grupo 1 engloba as espécies do subgênero Parviflorae (Lindley) Withner l990, colocando à parte duas espécies (Laelia harpophylla e Laelia kautskyi) que ficam excluídas, enquanto que a Laelia brevicaulis, considerada geralmente como uma espécie-irmã das duas precedentes, permanece. Entretanto, estas três espécies são, por sua vez, bem semelhantes entre elas e muito diferentes dos outros membros do subgênero Parviflorae; por outro lado, pode ter ocorrido um erro na amostragem de Laelia brevicaulis. Efetivamente, Laelia cowani Rolfe é dada (Withner, l990, Zappi, l995) como sinônimo de Laelia brevicaulis, enquanto que, na realidade, este nome é sinônimo de Laelia crispata; pode ter havido uma confusão no espécime analisado por Van den Berg et al. (C 208-Machado sn) e pode ter sido, na realidade, um espécime de Laelia crispata o que explicaria seu posicionamento quanto à proximidade imediata desta última espécie na árvore filogenética apresentada. Um estudo mais aprofundado do taxon Laelia brevicaulis é, pois, necessário e, por enquanto, nós preferimos adotar uma posição conservadora e colocar as três espécies, Laelia brevicaulis, Laelia harpophylla e Laelia kautskyi no grupo três, que coincide assim com a seção Harpophyllae Withner l990. Feita esta reserva, o resultados de van den Berg et al. confirmam que o subgênero Parviflorae, retirado da seção Harpophyllae, forma um grupo homogêneo e claramente distinto das outras espécies de Laelia. O grupo dois abrange as espécies de Laelia das seções Crispa Pftizer l889 (ou seção Cattleyodes Schlechter l917) e Hadrolaelia Schlecther l917, as espécies unifloras de Sophronitis , assim como a Cattleya máxima. Como reconhecem os próprios autores, o caso desta última espécie é suficientemente inesperado para merecer análises de verificação. Como eles, nós pensamos que é mais prudente esperar os resultados complementares antes de se pronunciar sobre seu status. Independentemente dos resultados, o grupo constitui uma outra entidade claramente delimitada. Efetivamente, do ponto de vista morfológico, Sophronitis coccinea, Sophronitis brevipedunculata (inicialmente descrita como uma variedade da Sophronitis wittigiana), Sophronitis mantiqueirae e Sophronitis wittigiana, apresentando todas uma inflorescência sem espata e uniflora (raramente biflora), são muito mais próxima das espécies da seção Hadrolaelia do que Sophronitis cernua, cuja inflorescência carrega diversas flores de pequeno tamanho. A este respeito, nos parece importante lembrar, que Porto e Brade tinham, desde l935, apontado que os dois grupos de Sophronitis apresentam diferenças morfológicas maiores: isto os levou a criar o gênero Eunannos para acolher as espécies unifloras (Porto & Brade, l935). A árvore filogenética de van den Berg et al. (op. cit.) parece, certamente, mostrar que este sub-grupo de Sophronitis é parafilético, mas, por outro lado, esta parte da árvore apresenta apenas uma fraca confiabilidade. O grupo 4 engloba apenas uma espécie, Sophronitis cernua. Esta espécie possui uma inflorescência multiflora que a distingue das outras espécies de Sophronitis colocadas no grupo 2. Como se trata também da espécie-tipo do gênero Sophronitis, o grupo 4 deve estar assimilado a este gênero. Teria sido interessante saber qual teria sido o lugar de Sophronitis pterocarpa, outra espécie multiflora do gênero. Enfim, Laelia lundii (grupo 5) ocupa, na árvore mais parcimoniosamente apresentada, uma posição isolada, de acordo com a concepção de Withner (op.cit.) que colocava esta espécie dentro de um subgênero especialmente criado para ela, o subgênero Microlaelia (Schlechter) Withner l990. Efetivamente, em função de suas características tanto morfológicas (2) quanto ecológicas (área de distribuição centrada ao nordeste de São Paulo e disjunta daquelas de outros grupos), esta espécie é, desde há muito tempo, tida como aberração dentro do gênero Laelia. Em sua discussão, van den Berg e seus co-autores indicam que uma solução alternativa à integração das espécies brasileiras de Laelia dentro do gênero Sophronitis seria de limitar, ao contrário, este gênero a 3 espécies, de separar Laelia lundii e de ressuscitar o gênero Hoffmannseggella para colocar nele as outras espécies brasileiras de Laelia e as outras espécies de Sophronitis. No entanto, eles consideram que a primeira solução seria preferível porque, segundo eles, não há grande diferença morfológica no interior do conjunto estudado. Justamente ao contrário, como demonstrou todos os estudos precedentes e como está preciso neste, as espécies abrangidas apresentam diferenças notáveis tanto em seus caracteres morfológicos quanto sobre o aspecto ecológico. Também, à luz dos comentários que acabam de ser feitos, nos parece mais adequado admitir que os resultados de van den Berg et al. reforçam as posições adotadas pelos botânicos acima citados e prosseguir com o trabalho destes últimos na delimitação das entidades autônomas. Assim,
nós propomos aqui considerar cada um dos grupos acima citados como
um gênero próprio, de assimilar o grupo 1 ao gênero
Hoffmannseggella Jones e de criar três novos gêneros
para organizar as espécies dos grupos 2, 3 e 5 (o grupo 4 ficando
assimilado ao gênero Sophronitis ): o subgênero Microlaelia
(grupo 5) está assim elevado à categoria de gênero;
A seção Harpophylla (grupo 3) fica transferida para
o gênero que dedicamos à Fritz Dungs; os três conjuntos
que constituem o grupo 2 ficam transferidos para um último gênero
cujo nome é retirado da seção Hadrolaelia.
Esta entidade reagrupa, na verdade, plantas epífitas relativamente
próximas, ainda que elas apresentem particularidades que permitem
reparti-las em diversas seções, como pode ser visto abaixo
(3). |
| In Acta Botanica Academiae Scientiarum Hungaricae 14(12) : 63-70 [69]. | |
| Jones
assim define o gênero: "Gênero diferente de Laelia em razão de seu porte e de um labelo muito mais estreito. Planta litófita ou raramente epífita de origem brasileira. Sépalas de dimensão igual às pétalas, ligeiramente mais curtas. 8 políneas". Em razão da exclusão das espécies transferidas para o gênero Dungsia (ver também § 3 sobre este gênero), é conveniente limitar este gênero às espécies rupícolas. De mais a mais, todos os representantes do gênero assim definido possuem pseudobulbos claviformes e unifoliados, inflorescências mais altas do que as folhas e flores pequenas (diâmetro inferior a 3cm). |
|
Sinonímia Laelia Lindley seção Parviflorae Lindley 1842 Laelia Lindley sub-seção Crispatae Cogniaux l901, in part. Laelia Lindley seção Cyrtolaelia Schlecther l917, in part. Laelia Lindley subgênero Parviflorae (Lindley) Withner l990, retirada da seção Harpophyla Withner l990 espécie-tipo: Hoffmannseggella cinnabarina (Bateman) H.G. Jones l968 basiônimo: Laelia cinnabarina Bateman ex Lindley l838, in Sert. Orch. t. 28 Withner (l990) organizava seu subgênero Parviflorae em 5 seções: Além da seção Harpophyla já citada, ele propunha separar as |
|
espécies rupícolas em 4 seções baseadas no comprimento
da inflorescência e na cor das flores. Estes caracteres não nos parecem critérios suficientes para delimitar as seções e, aqui, nós não nos prendemos a este conceito. (3) À vista da figura 1, nós poderíamos ficar tentados a dizer que os grupos 3 e 4 assim como os grupos 1 e 2, têm um ancestral comum e que estes dois ancestrais, por sua vez, têm eles mesmos um ancestral comum. E de propor uma organização em 3 gêneros sendo 2 divididos em 2 sub-gêneros. Entretanto, por todas as razões acima evocadas, esta conclusão seria prematura, ainda mais que ele não é sustentada de maneira evidente pelo estudo dos caracteres morfológicos |
|
basiônimo:
Laelia Lindley section Microlaelia Schlecther l917, in Orchids
11(5):87-96 [90]. Diagnosis - Herbae apiphyticae parvae, repentes, rhizomate elongato et pseudobulbis fusiformibus bifoliatisque; folia semiteretia, linearia; inflorescentia non orta ex spatha: pedunculs brevis sine nodis; petala sepalaque subsimilia; labellum tribolatum, cum 5 cristis longitudinalibus; 8 pollinia. Sinonímia Laelia Lindley subgênero Microlaelia (Schlecter) Withner l990. espécie-tipo: Microlaelia lundii (Rchb. f.) Chiron & V. P. Castro in Richardiana II (1): 11. 2002 basiônimo: Laelia lundi Rchb. f. l881, in Otia Bot Hamburg. 2;92 |
| Diagnosis - Herbae epiphyticae, rhizomate breviusculo et pseudobulbis cylindraceis, tenuibus, unifoliatis; folia elongata, anguste ligulata, coriacea; inflorescentia orta ex spatha, foliis multo brevior; flores mediocres, lutei, aurantiaci vel rubri; petala sepalaque subsimilia, labellum profunde trilobatum, lobo |
|
medio ondulatimarginato, deorsum curvato retroflexoque; 8 pollinia. Sinonímia Laelia Lindley sub-seção Harpophyllae Brieger l981, nomen nudum Laelia Lindley seção Harpophyllae Withner l990 espécie-tipo: Dungsia harpophylla (Rchb. f) Chiron & V. P. Castro in Richardiana II (1): 11. 2002 basiônimo: Laelia harpophylla Rchb. f. l873, in Gard. Chron. 542 |
|
basiônimo:
Laelia Lindley seção Hadrolaelia Schlechter
l917, in Orchids 11 (5):87-96 [89]. Diagnosis: Herbae epiphyticae pseudobulbis fusiformis, plus minusve elongatis, plus minusve longitudinaliter applanatis, unifoliatis; folium acaule, ellipticum vel ovatum, plus minusve coriaceum; flores persaepe magni, petalis sepalis latioribus, labelllo plus minusve trilobato, sinibus inter lobos laterales et lobum medium latis; 8 pollinia. espécie-tipo: Hadrolaelia pumila (Hooker) Chiron & V. P. Castro in Richardiana II (1): 11.2002. basiônimo: Cattleya pumila Hooker l838, in Bot. Mag. 65 t. 3656. |
|
|