|
| Keys word: Dungsia, Hadrolaelia, Hoffmannseggella, Laelia, Microlaelia , Brazil, Taxonomy. |
| The
genus Laelia has been divided for a long time in several entities
by different authors due to morphological observations. The existence of
two well-separated groups is confirmed by a recent study based on molecular
analysis of internal transcribed spacers (ITS) of nuclear ribosomal DNA:
Brazilian species should be removed from Mexican species (the true Laelia).
A strong congruence between morphological, environmental and molecular characteristics leads to resurrect the genus Hoffmannseggella H.G. Jones in order to accommodate rupicolous species; to put Laelia lundi Rchb. f., a very different species, in its own genus and to establish two other genera for the remaining species. Appropriate nothogenera are founded and new combinations proposed. |
| When
the genus Laelia has been created, in 1831, J. Lindley described
it as close to the genus Cattleya, but having 8 pollinia instead
of 4 and has founded this genus on two Mexican species early described as
belonged to the genus Bletia by La Llave & Lexarza, Bletia
grandiglora e Bletia autumnalis, excluding a Brazilian species,
nowadays known as Laelia crispa, putting it into the genus
Cattleya. Laelia are epiphytic or lithophyte plants, native of Mexico (and north of Central America) and Brazil. The pseudobulbs are swollen, more or less hollowed (some Mexican species), claviform, fusiform or cylindrical, having in the apex, one or two (occasionally even more) leaves which are, in general, leathery. The inflorescence is terminal and bears, sometimes, only one flower, sometimes many flowers, arranged in raceme or, more rarely, in panicle. Except for the rupícolas species, the flowers are, in general, big and showy and the color varies a lot. The petals and sepals are free, well-opened, spread, flattened or undulated. The lip is free or slightly adnate at the base of the column, it can be entire or trilobate, forming a tube around the column at the base and spreading afterward, the disc of the lip is flat or in laminas. The column is alongate and has a tooth in the apex. The apiculate anther shelters 8 waxy pollinia that are ovoid or compressed, disposed in 4 per each cellule. Soon after to have established the genus, Lindley has proposed the first sub-division and has particularly recognized a Brazilian section Parvilflorae (Lindley, 1842). After that, many authors (Cogniaux, 1901, Schlechter, 1917; Pabst & Dungs, 1975) have pointed out the heterogeneous character of the genus Laelia senso lato, re-grouping the Mexican species and the Brazilian species. Brieger (1981) advanced a step and divided the genus into two sub-genus: the sub-genus Eulaelia, which comprehends the Mexican species, the " true " Laelia since the type species (1) of the genus, Laelia speciosa (Kunth) Schlecther, is part of it and the sub-genus Brasiliensis, which gathers the Brazilian species. In fact, since many time ago, the orchidologists as well as the orchidists have been considering that the Mexican species of Laelia are much closer to Schomburgkia Lindley and to Encyclia Hook Hook than the Brazilian species of Laelia. On the other hand, in Brazil, Laelia purpurata Lindley is considered the most representative of the Brazilian group and under the point of view of the vegetative habit, it resembles much more to a Cattleya Lindley than to a Mexican Laelia. Even inside the Brazilian species, the morphological differences are big and more homogenous groups have been delimitated. So, H. G Jones (1968) created the genus Hoffmannseggella to the rupicolous species, however this conception was not followed by the taxonomists. Carrying on the classification of the genus, Withner proposed, some times ago, the creation of 3 sub-genera to organize the Brazilian species of Laelia (Withner, 1990); one of those sub-genera coincides, of course, with the genus Hoffmmannseggella H.G. Jones. But, may be to avoid a tumult in the milieu horticultural, Withner has never included the Brazilian species in this distinct genus. More recently, Halbinger and Soto presented a cladistic study about the relation between the members of what is called Laelia-Cattleya alliance founded on 64 morphological characters (Halbinger & Soto, 1997). This analysis confirms the distance between the Mexican species and the Brazilian species and the existence of natural group "Laelia subgenus Parviflorae" (with a bootstrap of 97%), however there is a lack of result concerning the groups "Laelia subgenus Crispae" and "Sophronitis ". All this taxonomical work, firstly founded just on the morphological characters of the plants, considers, nowadays, other characters, intrinsic (scent, blooming season) as well the extrinsic (pollinators, biogeography...). The recent evolution of the techniques of molecular analysis, allowing the access to the sequential of ADN parts, has considerably multiplied the number of the available characters. Recently, van den Berg and his co-authors have study the phylogeny of the sub-tribe Laeliinae through the molecular analysis of the sequence data internal transcribed spacers (ITS) of nuclear ribosomal ADN (van den Berg et al. 2000). This study confirms the existence, inside the genus Laelia sensu lato, of two ensembles very distant concerning the phylogenetic plan. Morphological, ecological and molecular data are, so, in harmony and this congruence reinforces the need for separating Brazilian species from the genus Laelia. van den Berg et al. were, therefore, the firsts to take this step. The phylogenetic tree related to Brazilian species of Laelia shows clearly the existence of many well delimitated clades (van den Berg et al. op. cit. fig 6, page 109), as we can see more evidenced in the figure 1 that reproduces the tree in a condensed way. The results brought van den Berg and his co-authors to re-group all Brazilian species of Laelia inside the genus Sophronitis. However, this conception brings, on one hand, to gather inside the same entity, plants which are morphologically very different and, on the other hand, to ignore the results of the predecessors: it seems to us to be necessary to discuss it and to propose an organization closer to the classical conception. (1) The name Bletia grandiflora La Llave & Lex. 1825, based on what the type species - Laelia grandiflora (La Llave & Lex) Lindley has been described, is, as a matter of fact, a synonym of Bletia speciosa Kinth, early published (1815). The last epithet has, thus, the priority. |
|
||
|
Figure
1: Cladogramme of Brazilian Laelias
(adapted from van der Berg et al., 2000) |
||
| The number above each branch (Fitch width) characterizes the number of changes noticed in the studied sequence along this branch. | ||
| |
| Before to move forward in the discussion of the results presented by those authors, we should remember some observations of general order about the own nature of the instrument "molecular phylogeny", observations that should bring us to have a great prudence regarding to the discussion of the results. |
| the study of the phylogenetic relations based on a particular gene analyzed inside an ensemble of species informs us, with a great rigor, about the phylogeny of this gene and not about the phylogeny of the species involved; this problem has been and still is avidly discussing (see mainly the well complete presentation done by Brower et al. 1996); only after obtaining concordant results about many genes, we can affirm, without running a great risk, that the phylogenetic tree thus established can be also applied to the studies species; |
| the molecular characters of the studied gene do not give an absolute and incontestable result (and, under this point of view, the sequence from ITS, in general used by reason of experimental convenience, is particularly weak); even if, no doubt about, less subjective than a studied based on the selection of morphological characters, the method presents its own limits and does not exempt an attentive studied about its characters; on the contrary, it should be used as a guide to retake the morphological analysis in a renewed optic, in order to solve eventual conflicts between the molecular data and morphological data (see, for example, Lecointre & Le Guyader 's discussion, 2001); |
| the analyzed sequence presents a low level of variation from one species to another, mainly in the interior of each one of the groups which are schematized in the figure 1; as admitted by the authors; "the weak levels of divergence could implicate in a sampling error as a reason of unexpected collocations". Any way, this characteristic brings a weak resolution in the extremity of the branches and does not allow an acute position of the species regarding to others in the interior of one group. We should notice, on the other hand, that the variations registered between two specimens of the same species (as happens, for example, with Cattleya maxima, Laelia xanthina or Sophronitis cernua) are, sometimes, more important than those which separate distinct species. |
| We
notice, in a small number of ramifications, very high frequencies of changes
(parallel development) such in Cattleya maxima, Laelia fidelensis,
Laelia virens, Laelia perrinii and Sophronitis cernua;
we noticed (this is the phenomenon known as attraction of the long ramifications)
that, when the velocities of evolution between the lineages are very different
one from the others, the species that evolves faster has more opportunity
to share the states of character by homoplasy than by ascendance; as a result,
the species that have long ramifications (it means, a great number of variation),
can be re-grouped together inside phylogenetic tree regardless of their
kinship. (2) We should notice that the global indexes of homoplasy for the sub-tribe, in the whole, are relativity important, with a coherence index of 0,26 and a retention index of 0,71. |
| certain clades are weakly supported by tests of coherence and robustness (percentage of bootstrap little superior and, often, inferior to 50%); still there, as even the authors point out, in those conditions, we should wait for the complementary results. |
|
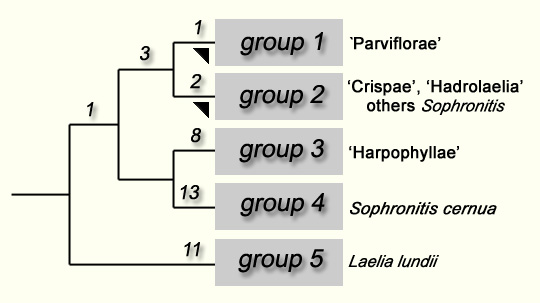
The group 1 comprehends the species of the sub-genus Parviflorae (Lindley) Withner 1990, putting aside two species (Laelia harpophylla and Laelia kautskyi) that stay excluded, while Laelia brevicaulis, in general considered as very close related to the two precedents, remains. However, those three species are very similar between them and very different from the other species of the sub-genus Parviflorae; On the other hand, it could have a mistake in the analyzed specimen of Laelia brevicaulis. In fact, Laelia cowani Rolfe is considered (Withner, 1990, Zappi, 1995) as a synonym of Laelia brevicaulis, however, as a matter of fact, this name is a synonym of Laelia crispata; it could have been a misunderstanding with the specimen analyzed by van den Berg et al. (C 208-Machado sn) and it could have been, in fact, a specimen of Laelia crispata that would explaine his opinion concerning the proximity of the last species in the phylogenetic tree presented. A deepened study of the taxon Laelia brevicaulis é is, therefore, necessary and, meanwhile, we prefer to adopt a conservative position and re-group the three species; Laelia brevicaulis, Laelia harpophylla and Laelia kautskyi in the group three which coincides with the section Harpophyllae Withner 1990. This reserve done, the results obtained by van den Berg et al. confirm that the sub-genus Parviflorae, removed from the section Harpophyllae, forms a homogenous group, clearly distinct from the other species of Laelia. The group two comprehends the species of Laelia from the sections Crispae Pftizer 1889 (or section Cattleyodes Schlechter 1917) and Hadrolaelia Schlecther 1917, the uniflora species of Sophronitis, as well Cattleya máxima. As recognized by the authors themselves, the event of this last species is enough unexpected to deserve an analysis of verification. As them, we think it is more prudent to wait the complementary results before pronouncing about its status. Regardless of the results, the group forms another entity clearly delimitated. Actually, under the morphological point of view, Sophronitis coccinea, Sophronitis brevipedunculata (firstly described as a variety of Sophronitis wittigiana), Sophronitis mantiqueirae and Sophronitis wittigiana, all of them presenting an uniflora (rarely bifloral) inflorescence without sheath, are closer to the species of the section Hadrolaelia than the Sophronitis cernua, which inflorescence bears many small flowers. It seems important to remember that, concerning this subject, Porto and Brade had, since 1935, pointed out that the two groups of Sophronitis present biggest morphological differences: this brought them to create the genus Eunannos to place the uniflora species (Porto & Brade, 1935). The phylogenetic tree established by van den Berg et al. (op. cit.) seems, certainly, show that this sub-group of Sophronitis is paraphyletic, however, on the other hand, this part of the tree presents just a weak reliably. The group 4 comprehends just one species, Sophronitis cernua. This species have a multiflora inflorescence that distinguishes it from the other species of Sophronitis placed in the group 2. As it is also the type species of the genus Sophronitis, the group 4 should be placed in this genus. It would be very interesting to know which would be the place of Sophronitis pterocarpa, another multifora species of the genus. Finally, Laelia lundii (group 5) occupies, in the more parsimonious tree presented, an isolated position, according to Withner's conception (op.cit.) who put this species inside a sub-genus specially created to it, Microlaelia (Schlechter) Withner 1990. In fact, regarding its morphological characteristics (see §2 bellow) as well the ecological ones (with the center of the distribution area located in the north-east of São Paulo and disjoints of those from the other groups), this species is considered, since long time ago, as an aberration inside the genus Laelia. In his discussion, van den Berg and his co-authors show that the alternative solution to the integration of Brazilian species of Laelia inside the genus Sophronitis would be to limit, on the contrary, this genus into 3 species, to separate Laelia lundii and to ressurect the genus Hoffmannseggella to put inside it the other Brazilian species of Laelia and the other species of Sophronitis. However, they consider that the first solution would be preferable because, according to them, there is no a great morphological difference inside the studied group. Quite the contrary, as showed by all precedent studies and as specified in this one, the included species present remarkable differences in their morphological characters as well in their ecological aspect. Besides, in the light of the commentaries that have just been done, it seems to us more adequate to admit that the results obtained by van den Berg et al. reinforce the positions adopted by the botanists above mentioned and to carry on with their work in the delimitation of the autonomy entities. So, we propose here to consider every one of those groups as an own genus, to include the group 1 to the genus Hoffmannseggella Jones and to create three new genus to organize the species of the groups 2, 3 e 5 (the group 4 included in the genus Sophronitis ): the subgenus Microlaelia (group 5) is brought the category of genus; The section Harpophylla (group 3) is transferred to the genus named in honor of Fritiz Dungs; the three ensembles which form the group 2 are transferred to a last genus which name is taken from the section Hadrolaelia. This entity re-groups, in fact, epiphyte plants relatively close, even though they present particularities that allow to divide them into many sections as showed bellow (also see the note (3). |
| In Acta Botanica Academiae Scientiarum Hungaricae 14(12) : 63-70 [69]. | |
| Jones
defines the genus: "Genus different from Laelia by size and by a much more narrow lip. Lithophyte or rarely epiphytic plants from Brazil. Sepals with the same size of petals, just a little shorter. 8 pollinia". Due the exclusion of the species transferred to the genus Dungsia (see § 3 below), it is convienient to limit this genus to the rocky species. Anyhow, all members of the genus defined in this way have claviform and unifoliate pseudobulbs, and floral stalks higher than the leaves and small flowers (diameter less than 3cm). |
|
Synonymy: Laelia Lindley section Parviflorae Lindley 1842 Laelia Lindley sub-section Crispatae Cogniaux 1901, in part. Laelia Lindley section Cyrtolaelia Schlecther 1917, in part. Laelia Lindley subgenus Parviflorae (Lindley) Withner 1990, removed from the section Harpophylae Withner 1990 Type species: Hoffmannseggella cinnabarina (Bateman) H.G. Jones 1968 Basionym: Laelia cinnabarina Bateman ex Lindley 1838, in Sert. Orch. t. 28 Withner (1990) organized the subgenus Parviflorae into 5 sections: Besides the section Harpophylae as already mentioned, he proposed to separate the rupicolous species |
|
into 4 sections based on the length of the inflorescence and on the flowers
color. Those characters do not seem to us enough to delimitate the sections
and, here, we do not retain this concept. (3) Examining the figure 1, we could be inclined to say that the groups 3 and 4 have a common ancestral as well the groups 1 and 2 and that those two ancestors have a common ancestor. And to suggest the organization into 3 genera where 2 would be divided into 2 sub-genera. However, due the reasons referred above, this conclusion would be premature moreover it is not sustained in an evident way by the study of morphological characters. |
|
Basionym:
Laelia Lindley section Microlaelia Schlecther 1917, in Orchids
11(5):87-96 [90]. Diagnosis - Herbae apiphyticae parvae, repentes, rhizomate elongato et pseudobulbis fusiformibus bifoliatisque; folia semiteretia, linearia; inflorescentia non orta ex spatha: pedunculs brevis sine nodis; petala sepalaque subsimilia; labellum tribolatum, cum 5 cristis longitudinalibus; 8 pollinia. Sinonymy Laelia Lindley subgenus Microlaelia (Schlecter) Withner 1990. Type species: Microlaelia lundii (Rchb. f.) Chiron & V. P. Castro in Richardiana II (1): 11. 2002. Basionym: Laelia lundi Rchb. f. 1881, in Otia Bot Hamburg. 2;92 |
| Diagnosis - Herbae epiphyticae, rhizomate breviusculo et pseudobulbis cylindraceis, tenuibus, unifoliatis; folia elongata, anguste ligulata, coriacea; inflorescentia orta ex spatha, foliis multo brevior; flores mediocres, lutei, aurantiaci vel rubri; petala sepalaque subsimilia, labellum profunde trilobatum, lobo |
|
medio ondulatimarginato, deorsum curvato retroflexoque; 8 pollinia. Synonymy Laelia Lindley sub-section Harpophyllae Brieger 1981, nomen nudum Laelia Lindley section Harpophyllae Withner 1990. Type species: Dungsia harpophylla (Rchb. f) Chiron & V. P. Castro in Richardiana II (1): 11. .2002. Basionym: Laelia harpophylla Rchb. f. 1873, in Gard. Chron. 542. |
|
Basionym:
Laelia Lindley section Hadrolaelia Schlechter 1917, in Orchids
11 (5):87-96 [89]. Diagnosis: Herbae epiphyticae pseudobulbis fusiformis, plus minusve elongatis, plus minusve longitudinaliter applanatis, unifoliatis; folium acaule, ellipticum vel ovatum, plus minusve coriaceum; flores persaepe magni, petalis sepalis latioribus, labelllo plus minusve trilobato, sinibus inter lobos laterales et lobum medium latis; 8 pollinia. Type species: Hadrolaelia pumila (Hooker) Chiron & V. P. Castro in Richardiana II (1): 11.2002. basionym: Cattleya pumila Hooker 1838, in Bot. Mag. 65 t. 3656. |
|
|